The intensive maize-wheat double-cropping system underpins food security in China's Huang-Huai-Hai Plain but drives soil degradation through nutrient depletion and biodiversity loss, necessitating sustainable diversification. This study mechanistically compared legacy effects of five preceding summer crops-maize (control), soybean, peanut, pepper, sweet potato-on subsequent winter wheat performance, explicitly quantifying impacts on root architecture, soil enzymatic activity, and yield formation. Results demonstrated peanut’s unparalleled efficacy: it increased seedling-stage wheat shoot biomass by 37-41% and root biomass by 184% versus maize, while expanding root surface area (51%) and volume (54%) through optimized rhizosphere engineering. These morphological advantages persisted through maturity and correlated with significantly enhanced soil functionality-peanut elevated soil organic matter (25-37%), nitrate-N (138-148%), and ammonium-N (71-128%) while reducing C:N ratio. Crucially, peanut residues stimulated microbial metabolism, increasing β-glucosidase activity (governing C cycling) by 33-89% and urease activity (N mineralization) by 40-109%, whereas catalase activity showed context-dependent responses. This accelerated nutrient mineralization translated to agronomic superiority: peanut-wheat rotation yielded 10.5% more grain than maize-wheat, exceeding soybean-wheat by 3.4% despite lower 1000-grain weight, primarily through 26.6% higher ear density. Soybean provided intermediate soil N benefits but weaker root stimulation, while pepper suppressed enzymes and sweet potato inconsistently affected fertility. We conclude that peanut’s unique residue composition-low C:N ratio, abundant labile carbon, and rhizodeposits-primes a self-reinforcing root-microbe-soil loop that enhances nutrient synchrony, breaks maize-wheat yield ceilings, and offers a validated pathway for ecological intensification in cereal systems.
| Published in | Advances in Applied Sciences (Volume 10, Issue 3) |
| DOI | 10.11648/j.aas.20251003.14 |
| Page(s) | 74-87 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2025. Published by Science Publishing Group |
Crop Rotation, Preceding Crop Legacy, Root System, Soil Fertility, Soil Enzyme Activity, Peanut-wheat System
RCBD | Randomized Complete Block Design |
ANOVA | Rnalysis Of Variance |
LSD | Least Significant Difference |
| [1] | Shi, W.; Wang, M.; Tao, F.; Xu, X.; Deng, X.; Liu, L.; Kong, X.; Zuo, L.; Lei, M.; Shi, X.; et al. Wheat Redistribution in Huang-Huai-Hai, China, Could Reduce Groundwater Depletion and Environmental Footprints without Compromising Production. Commun Earth Environ 2024, 5, 380, |
| [2] | Yin, L.; Tao, F.; Chen, Y.; Wang, Y.; Ciais, P.; Smith, P. Novel Cropping-System Strategies in China Can Increase Plant Protein with Higher Economic Value but Lower Greenhouse Gas Emissions and Water Use. One Earth 2023, 6, 560-572, |
| [3] | ZHANG Quan-Jun; WU Dong-Li; Jing, G. Variation of Winter Wheat Phenology Dataset in Huang Huai Hai Plain of China from 1981 to 2021 Variation of Winter Wheat Phenology Dataset in Huang Huai Hai Plain of China from 1981 to 2021 2025, 1631413680 bytes, 1 files. |
| [4] | Gong, K.; Rong, L.; Zhang, Y.; Wang, X.; Duan, F.; Li, X.; He, Z.; Jiang, T.; Chen, S.; Feng, H.; et al. Efficient Agronomic Practices Narrow Yield Gaps and Alleviate Climate Change Impacts on Winter Wheat Production in China. Commun Earth Environ 2025, 6, 290, |
| [5] | Liang, Q.; Chen, H.; Gong, Y.; Fan, M.; Yang, H.; Lal, R.; Kuzyakov, Y. Effects of 15 Years of Manure and Inorganic Fertilizers on Soil Organic Carbon Fractions in a Wheat-Maize System in the North China Plain. Nutr Cycl Agroecosyst 2012, 92, 21-33, |
| [6] | Yang, J.; Zhang, S.; Zhang, J.; Zhao, S.; Lu, H.; Li, L.; Liu, L.; Wang, G. Incorporating Crop Rotation into the Winter Wheat-Summer Maize System to Enhance Soil Multifunctionality and Sustainable Grain Production in the North China Plain. Field Crops Research 2025, 325, 109834, |
| [7] | Grote, U.; Fasse, A.; Nguyen, T. T.; Erenstein, O. Food Security and the Dynamics of Wheat and Maize Value Chains in Africa and Asia. Front. Sustain. Food Syst. 2021, 4, 617009, |
| [8] | Landschoot, S.; Audenaert, K.; Waegeman, W.; De Baets, B.; Haesaert, G. Influence of Maize-Wheat Rotation Systems on Fusarium Head Blight Infection and Deoxynivalenol Content in Wheat under Low versus High Disease Pressure. Crop Protection 2013, 52, 14-21, |
| [9] | Tamburini, G.; Bommarco, R.; Wanger, T. C.; Kremen, C.; Van Der Heijden, M. G. A.; Liebman, M.; Hallin, S. Agricultural Diversification Promotes Multiple Ecosystem Services without Compromising Yield. Sci. Adv. 2020, 6, eaba1715, |
| [10] | Zani, C. F.; Manning, D. A. C.; Abbott, G. D.; Taylor, J. A.; Cooper, J.; Lopez-Capel, E. Diversified Crop Rotations and Organic Amendments as Strategies for Increasing Soil Carbon Storage and Stabilisation in UK Arable Systems. Front. Environ. Sci. 2023, 11, 1113026, |
| [11] | Yang, X.; Xiong, J.; Du, T.; Ju, X.; Gan, Y.; Li, S.; Xia, L.; Shen, Y.; Pacenka, S.; Steenhuis, T. S.; et al. Diversifying Crop Rotation Increases Food Production, Reduces Net Greenhouse Gas Emissions and Improves Soil Health. Nat Commun 2024, 15, 198, |
| [12] | Liang, Z.; Xu, Z.; Cheng, J.; Ma, B.; Cong, W.-F.; Zhang, C.; Zhang, F.; Van Der Werf, W.; Groot, J. C. J. Designing Diversified Crop Rotations to Advance Sustainability: A Method and an Application. Sustainable Production and Consumption 2023, 40, 532-544, |
| [13] | Yang, N.; Wang, Z.; Gao, Y.; Zhao, H.; Li, K.; Li, F.; Malhi, S. S. Effects of Planting Soybean in Summer Fallow on Wheat Grain Yield, Total N and Zn in Grain and Available N and Zn in Soil on the Loess Plateau of China. European Journal of Agronomy 2014, 58, 63-72, |
| [14] | Zou, X.; Huang, M.; Liu, Y.; Si, T.; Zhang, X.; Yu, X.; Guo, F.; Wan, S. Inclusion of Peanut in Wheat-Maize Rotation Increases Wheat Yield and Net Return and Improves Soil Organic Carbon Pool by Optimizing Bacterial Community. Journal of Integrative Agriculture 2023, 22, 3430-3443, |
| [15] | Zhang, K.; Wang, X.; Li, Y.; Zhao, J.; Yang, Y.; Zang, H.; Zeng, Z. Peanut Residue Incorporation Benefits Crop Yield, Nitrogen Yield, and Water Use Efficiency of Summer Peanut - Winter Wheat Systems. Field Crops Research 2022, 279, 108463, |
| [16] | Navarro, J.; Salazar, J.; Kang, J. J.; Parsons, J.; Cheng, C.-L.; Castillo, A.; Pujol Pereira, E. I. Compost and Biochar to Promote Soil Biological Activities under Sweet Potatoes Cultivation in a Subtropical Semiarid Region. Applied and Environmental Soil Science 2020, 2020, 1-11, |
| [17] | Shen, S.; Xu, G.; Clements, D. R.; Jin, G.; Chen, A.; Zhang, F.; Kato-Noguchi, H. Suppression of the Invasive Plant Mile-a-Minute (Mikania Micrantha) by Local Crop Sweet Potato (Ipomoea Batatas) by Means of Higher Growth Rate and Competition for Soil Nutrients. BMC Ecol 2015, 15, 1, |
| [18] | Younis, U.; Danish, S.; Datta, R.; Al Obaid, S.; Ansari, M. J. Synergistic Effects of Boron and Saponin in Mitigating Salinity Stress to Enhance Sweet Potato Growth. Sci Rep 2024, 14, 12988, |
| [19] | Yao, B.; Wang, X.; Li, Y.; Lian, J.; Li, Y.; Luo, Y.; Li, Y. Soil Extracellular Enzyme Activity Reflects the Change of Nitrogen to Phosphorus Limitation of Microorganisms during Vegetation Restoration in Semi-Arid Sandy Land of Northern China. Front. Environ. Sci. 2023, 11, 1298027, |
| [20] | Bowker, M. A.; Mau, R. L.; Maestre, F. T.; Escolar, C.; Castillo-Monroy, A. P. Functional Profiles Reveal Unique Ecological Roles of Various Biological Soil Crust Organisms: Functional Profiles of Biological Crusts. Functional Ecology 2011, 25, 787-795, |
| [21] | Bailey, V. L.; Fansler, S. J.; Stegen, J. C.; McCue, L. A. Linking Microbial Community Structure to β -Glucosidic Function in Soil Aggregates. The ISME Journal 2013, 7, 2044-2053, |
| [22] | Silva-Olaya, A. M.; Mora-Motta, D. A.; Cherubin, M. R.; Grados, D.; Somenahally, A.; Ortiz-Morea, F. A. Soil Enzyme Responses to Land Use Change in the Tropical Rainforest of the Colombian Amazon Region. PLoS ONE 2021, 16, e0255669, |
| [23] | Cao, X.; Shi, Z.; Chen, J.; Liu, S.; Zhang, M.; Chen, M.; Xu, G.; Wu, J.; Xing, H.; Li, F. Extracellular Enzyme Characteristics and Microbial Metabolic Limitation in Soil of Subalpine Forest Ecosystems on the Eastern Qinghai-Tibetan Plateau. Plant Soil 2022, 479, 337-353, |
| [24] | Daunoras, J.; Kačergius, A.; Gudiukaitė, R. Role of Soil Microbiota Enzymes in Soil Health and Activity Changes Depending on Climate Change and the Type of Soil Ecosystem. Biology 2024, 13, 85, |
| [25] | Uwituze, Y.; Nyiraneza, J.; Fraser, T. D.; Dessureaut-Rompré, J.; Ziadi, N.; Lafond, J. Carbon, Nitrogen, Phosphorus, and Extracellular Soil Enzyme Responses to Different Land Use. Front. Soil Sci. 2022, 2, 814554, |
| [26] | Asensio, D.; Zuccarini, P.; Sardans, J.; Marañón-Jiménez, S.; Mattana, S.; Ogaya, R.; Mu, Z.; Llusià, J.; Peñuelas, J. Soil Biomass-Related Enzyme Activity Indicates Minimal Functional Changes after 16 Years of Persistent Drought Treatment in a Mediterranean Holm Oak Forest. Soil Biology and Biochemistry 2024, 189, 109281, |
| [27] | Schaap, K. J.; Fuchslueger, L.; Quesada, C. A.; Hofhansl, F.; Valverde-Barrantes, O.; Camargo, P. B.; Hoosbeek, M. R. Seasonal Fluctuations of Extracellular Enzyme Activities Are Related to the Biogeochemical Cycling of C, N and P in a Tropical Terra-Firme Forest. Biogeochemistry 2023, 163, 1-15, |
| [28] | Zang, X.; Liu, M.; Fan, Y.; Xu, J.; Xu, X.; Li, H. The Structural and Functional Contributions of β-Glucosidase-Producing Microbial Communities to Cellulose Degradation in Composting. Biotechnol Biofuels 2018, 11, 51, |
| [29] | Bai, X.; Wang, X.; Wang, S.; Ji, X.; Guan, Z.; Zhang, W.; Lu, X. Functional Studies of β-Glucosidases of Cytophaga Hutchinsonii and Their Effects on Cellulose Degradation. Front. Microbiol. 2017, 8, |
| [30] | Zhang, X.; Ma, B.; Liu, J.; Chen, X.; Li, S.; Su, E.; Gao, L.; Li, H. β-Glucosidase Genes Differentially Expressed during Composting. Biotechnol Biofuels 2020, 13, 174, |
| [31] | Adetunji, A. T.; Ncube, B.; Meyer, A. H.; Olatunji, O. S.; Mulidzi, R.; Lewu, F. B. Soil pH, Nitrogen, Phosphatase and Urease Activities in Response to Cover Crop Species, Termination Stage and Termination Method. Heliyon 2021, 7, e05980, |
| [32] | Sant’Anna, S. A. C.; Martins, M. R.; Goulart, J. M.; Araújo, S. N.; Araújo, E. S.; Zaman, M.; Jantalia, C. P.; Alves, B. J. R.; Boddey, R. M.; Urquiaga, S. Biological Nitrogen Fixation and Soil N2O Emissions from Legume Residues in an Acrisol in SE Brazil. Geoderma Regional 2018, 15, e00196, |
| [33] | Allende-Montalbán, R.; Martín-Lammerding, D.; Delgado, M. D. M.; Porcel, M. A.; Gabriel, J. L. Urease Inhibitors Effects on the Nitrogen Use Efficiency in a Maize-Wheat Rotation with or without Water Deficit. Agriculture 2021, 11, 684, |
| [34] | Stpniewska, Z.; Wolińska, A.; Ziomek, J. Response of Soil Catalase Activity to Chromium Contamination. Journal of Environmental Sciences 2009, 21, 1142-1147, |
| [35] | Guwy, A. J.; Martin, S. R.; Hawkes, F. R.; Hawkes, D. L. Catalase Activity Measurements in Suspended Aerobic Biomass and Soil Samples. Enzyme and Microbial Technology 1999, 25, 669-676, |
| [36] | Liu, J.; Sun, B.; Shen, H.; Ding, P.; Ning, D.; Zhang, J.; Qiu, X. Crop Water Requirement and Utilization Efficiency-Based Planting Structure Optimization in the Southern Huang-Huai-Hai Plain. Agronomy 2022, 12, 2219, |
| [37] | Yan, H.; Shengyuan, Y.; Li, M.; Xiao, J. Evaluation of Heavy Metal Contamination in Agricultural Topsoils in Suburban Xuchang City, China. Environ Earth Sci 2015, 74, 2475-2480, |
| [38] | Fan, X.; Characters on Available Nutrients of Soils under Different Land Use Types in Xuchang. OJSWC 2015, 03, 65-74, |
| [39] | Yeomans, J. C.; Bremner, J. M. A Rapid and Precise Method for Routine Determination of Organic Carbon in Soil. Communications in Soil Science and Plant Analysis 1988, 19, 1467-1476, |
| [40] | Official Methods of Analysis of AOAC INTERNATIONAL; Latimer, G. W., Ed.; 22nd ed.; Oxford University Press: New York, 2023; ISBN 978-0-19-761013-8. |
| [41] | Ye, C.; Cheng, X.; Liu, W.; Zhang, Q. Revegetation Impacts Soil Nitrogen Dynamics in the Water Level Fluctuation Zone of the Three Gorges Reservoir, China. Science of The Total Environment 2015, 517, 76-85, |
| [42] | Descheemaeker, K.; Nyssen, J.; Rossi, J.; Poesen, J.; Haile, M.; Raes, D.; Muys, B.; Moeyersons, J.; Deckers, S. Sediment Deposition and Pedogenesis in Exclosures in the Tigray Highlands, Ethiopia. Geoderma 2006, 132, 291-314, |
| [43] | Pei, B.; Liu, T.; Xue, Z.; Cao, J.; Zhang, Y.; Yu, M.; Liu, E.; Xing, J.; Wang, F.; Ren, X.; et al. Effects of Biofertilizer on Yield and Quality of Crops and Properties of Soil Under Field Conditions in China: A Meta-Analysis. Agriculture 2025, 15, 1066, |
| [44] | Cao, H.; Li, X.; Han, H.; Chen, S.; Jin, J.; Yuan, J.; Liang, C.; Lu, J.; Cui, F.; Chen, J. Exploring the Potential of Trichoderma Asperellum TCS007 on Growth Promotion of Pecan Seedlings as Well as Rhizosphere Soil Nutrients and Microbial Community. Plant Soil 2025, |
| [45] | Hu, J.; Yu, W.; Liu, P.; Zhao, B.; Zhang, J.; Ren, B. Responses of Canopy Functionality, Crop Growth and Grain Yield of Summer Maize to Shading, Waterlogging, and Their Combination Stress at Different Crop Stages. European Journal of Agronomy 2023, 144, 126761, |
| [46] | Zhang, K.; Zhao, J.; Wang, X.; Xu, H.; Zang, H.; Liu, J.; Hu, Y.; Zeng, Z. Estimates on Nitrogen Uptake in the Subsequent Wheat by Above-Ground and Root Residue and Rhizodeposition of Using Peanut Labeled with 15N Isotope on the North China Plain. Journal of Integrative Agriculture 2019, 18, 571-579, |
| [47] | Jani, A. D.; Mulvaney, M. J.; Balkcom, K. S.; Wood, C. W.; Jordan, D. L.; Wood, B. H.; Devkota, P. Peanut Residues Supply Minimal Plant-available Nitrogen on a Major Soil Series in the USA Peanut Basin. Soil Use and Management 2020, 36, 274-284, |
| [48] | Amissah, S.; Baidoo, M.; Agyei, B. K.; Ankomah, G.; Black, R. A.; Perry, C. D.; Hollifield, S.; Kusi, N. Y.; Harris, G. H.; Sintim, H. Y. Early and Late Season Nutrient Stress Conditions: Impact on Cotton Productivity and Quality. Agronomy 2022, 13, 64, |
| [49] | Fall, D.; Bakhoum, N.; Fall, F.; Diouf, F.; Ndiaye, C.; Faye, M. N.; Hocher, V.; Diouf, D. Effect of Peanut Shells Amendment on Soil Properties and Growth of Seedlings of Senegalia Senegal (L.) Britton, Vachellia Seyal (Delile) P. Hurter, and Prosopis Juliflora (Swartz) DC in Salt-Affected Soils. Annals of Forest Science 2018, 75, 32, |
| [50] | Mulvaney, M. J.; Balkcom, K. S.; Wood, C. W.; Jordan, D. Peanut Residue Carbon and Nitrogen Mineralization under Simulated Conventional and Conservation Tillage. Agronomy Journal 2017, 109, 696-705, |
| [51] | Jani, A. D.; Mulvaney, M. J.; Enloe, H. A.; Erickson, J. E.; Leon, R. G.; Rowland, D. L.; Wood, C. W. Peanut Residue Distribution Gradients and Tillage Practices Determine Patterns of Nitrogen Mineralization. Nutr Cycl Agroecosyst 2019, 113, 63-76, |
| [52] | Ye, X.; Ye, Y.; Chai, R.; Li, J.; Ma, C.; Li, H.; Xiong, Q.; Gao, H. The Influence of a Year-Round Tillage and Residue Management Model on Soil N Fractions in a Wheat-Maize Cropping System in Central China. Sci Rep 2019, 9, 4767, |
| [53] | Kong, L. Maize Residues, Soil Quality, and Wheat Growth in China. A Review. Agron. Sustain. Dev. 2014, 34, 405-416, |
| [54] | Barus, J.; Lumbanraja, J.; Sudarsono, H. A Litterbag Study: Decomposition Rate and C/N Ratio of Annual Crop Biomass Residues on An Ultisols in Natar Village, South Lampung, Indonesia. TROPICAL AGRICULTURAL SCIENCE 2019, 42, 387-403. |
| [55] | Surigaoge, S.; Yang, H.; Su, Y.; Du, Y.-H.; Ren, S.-X.; Fornara, D.; Christie, P.; Zhang, W.-P.; Li, L. Maize/Peanut Intercropping Has Greater Synergistic Effects and Home-Field Advantages than Maize/Soybean on Straw Decomposition. Front. Plant Sci. 2023, 14, 1100842, |
| [56] | Han, L.; Zhang, H.; Bai, X.; Jiang, B. The Peanut Root Exudate Increases the Transport and Metabolism of Nutrients and Enhances the Plant Growth-Promoting Effects of Burkholderia Pyrrocinia Strain P10. BMC Microbiol 2023, 23, 85, |
| [57] | Ankati, S.; Rani, T. S.; Podile, A. R. Changes in Root Exudates and Root Proteins in Groundnut-Pseudomonas Sp. Interaction Contribute to Root Colonization by Bacteria and Defense Response of the Host. J Plant Growth Regul 2019, 38, 523-538, |
| [58] | Ma, L.; Ma, S.; Chen, G.; Lu, X.; Chai, Q.; Li, S. Mechanisms and Mitigation Strategies for the Occurrence of Continuous Cropping Obstacles of Legumes in China. Agronomy 2023, 14, 104, |
| [59] | De Camargo, A. C.; Regitano-d’Arce, M. A. B.; Rasera, G. B.; Canniatti-Brazaca, S. G.; Do Prado-Silva, L.; Alvarenga, V. O.; Sant’Ana, A. S.; Shahidi, F. Phenolic Acids and Flavonoids of Peanut By-Products: Antioxidant Capacity and Antimicrobial Effects. Food Chemistry 2017, 237, 538-544, |
| [60] | Mandal, S. M.; Chakraborty, D.; Dey, S. Phenolic Acids Act as Signaling Molecules in Plant-Microbe Symbioses. Plant Signaling & Behavior 2010, 5, 359-368, |
| [61] | Zhang, W.; Sun, K.; Shi, R.-H.; Yuan, J.; Wang, X.-J.; Dai, C.-C. Auxin Signalling of Arachis Hypogaea Activated by Colonization of Mutualistic Fungus Phomopsis Liquidambari Enhances Nodulation and N2 -Fixation: Auxin in Fungus-Legume-Rhizobium Interactions. Plant Cell Environ 2018, |
| [62] | Saini, N.; Anmol, A.; Kumar, S.; Wani, A. W.; Bakshi, M.; Dhiman, Z. Exploring Phenolic Compounds as Natural Stress Alleviators in Plants- a Comprehensive Review. Physiological and Molecular Plant Pathology 2024, 133, 102383, |
| [63] | Ali, S.; Liu, K.; Ahmed, W.; Jing, H.; Qaswar, M.; Kofi Anthonio, C.; Maitlo, A. A.; Lu, Z.; Liu, L.; Zhang, H. Nitrogen Mineralization, Soil Microbial Biomass and Extracellular Enzyme Activities Regulated by Long-Term N Fertilizer Inputs: A Comparison Study from Upland and Paddy Soils in a Red Soil Region of China. Agronomy 2021, 11, 2057, |
| [64] | Li, X.; Hou, L.; Liu, M.; Lin, X.; Li, Y.; Li, S. Primary Effects of Extracellular Enzyme Activity and Microbial Community on Carbon and Nitrogen Mineralization in Estuarine and Tidal Wetlands. Appl Microbiol Biotechnol 2015, 99, 2895-2909, |
| [65] | Zhang, L.; Jing, Y.; Chen, C.; Xiang, Y.; Rezaei Rashti, M.; Li, Y.; Deng, Q.; Zhang, R. Effects of Biochar Application on Soil Nitrogen Transformation, Microbial Functional Genes, Enzyme Activity, and Plant Nitrogen Uptake: A Meta-analysis of Field Studies. GCB Bioenergy 2021, 13, 1859-1873, |
| [66] | He, W.; Zhang, M.; Jin, G.; Sui, X.; Zhang, T.; Song, F. Effects of Nitrogen Deposition on Nitrogen-Mineralizing Enzyme Activity and Soil Microbial Community Structure in a Korean Pine Plantation. Microb Ecol 2021, 81, 410-424, |
| [67] | Huang, C.; Feng, Y.; Patel, G.; Xu, X.; Qian, J.; Liu, Q.; Kai, G. Production, Immobilization and Characterization of Beta-Glucosidase for Application in Cellulose Degradation from a Novel Aspergillus Versicolor. International Journal of Biological Macromolecules 2021, 177, 437-446, |
| [68] | Li, X.; Zhang, Y.; Zhou, C.; Li, X.; Zou, X.; Ou, L.; Tao, Y. The Changes of Rhizosphere Microbial Communities in Pepper Varieties with Different Capsaicinoids. Front. Microbiol. 2024, 15, 1430682, |
| [69] | Füchtbauer, S.; Mousavi, S.; Bereswill, S.; Heimesaat, M. M. Antibacterial Properties of Capsaicin and Its Derivatives and Their Potential to Fight Antibiotic Resistance - A Literature Survey. EuJMI 2021, 11, 10-17, |
| [70] | Du, X.; Zhang, X.; Kong, L.; Xi, M. Split Application under Reduced Nitrogen Rate Favors High Yield by Altering Endogenous Hormones and C/N Ratio in Sweet Potato. Agronomy 2020, 10, 1243, |
| [71] | Zhang, E.; Li, J.; Zhang, K.; Wang, F.; Yang, H.; Zhi, S.; Liu, G. Anaerobic Digestion Performance of Sweet Potato Vine and Animal Manure under Wet, Semi-Dry, and Dry Conditions. AMB Expr 2018, 8, 45, |
| [72] | Zhang, X.; Hua, Y.; Liu, Y.; He, M.; Ju, Z.; Dai, X. Wide Belt Sowing Improves the Grain Yield of Bread Wheat by Maintaining Grain Weight at the Backdrop of Increases in Spike Number. Front. Plant Sci. 2022, 13, 992772, |
| [73] | Nadia; Amanullah; Arif, M.; Muhammad, D. Improvement in Wheat Productivity with Integrated Management of Beneficial Microbes along with Organic and Inorganic Phosphorus Sources. Agriculture 2023, 13, 1118, |
| [74] | Xu, X.; Liu, S.; Meng, F.; Zhang, X.; Zhao, J.; Qu, W.; Shi, Y.; Zhao, C. Grain Yield Formation and Nitrogen Utilization Efficiency of Different Winter Wheat Varieties under Rainfed Conditions in the Huang-Huai-Hai Plain. Agronomy 2023, 13, 915, |
| [75] | Farhad, Md.; Tripathi, S.; Singh, R.; Joshi, A.; Bhati, P.; Vishwakarma, M.; Kumar, U. GWAS for Early-Establishment QTLs and Their Linkage to Major Phenology-Affecting Genes (Vrn, Ppd, and Eps) in Bread Wheat. Genes 2023, 14, 1507, |
APA Style
Hecheng, L., Xinru, H., Shancong, W., Ahmed, R. M. M., Abdelhadi, E. E. M., et al. (2025). Peanut Precursor Surpasses Soybean and Breaks the Maize-Wheat Paradigm by Engineering a Superior Rhizosphere to Boost Soil and Yield. Advances in Applied Sciences, 10(3), 74-87. https://doi.org/10.11648/j.aas.20251003.14
ACS Style
Hecheng, L.; Xinru, H.; Shancong, W.; Ahmed, R. M. M.; Abdelhadi, E. E. M., et al. Peanut Precursor Surpasses Soybean and Breaks the Maize-Wheat Paradigm by Engineering a Superior Rhizosphere to Boost Soil and Yield. Adv. Appl. Sci. 2025, 10(3), 74-87. doi: 10.11648/j.aas.20251003.14
@article{10.11648/j.aas.20251003.14,
author = {Liu Hecheng and Hu Xinru and Wang Shancong and Reda Mohamed Mahmoud Ahmed and Ebtesam Eid Mohamed Abdelhadi and Yu Qihang and Chen Meiyu and Meng Fanzheng and Wang Bo and Tang Yulou and Nasr Mahmoud Abdou and Kou Yanling and Zhang Rui and Shao Ruixin},
title = {Peanut Precursor Surpasses Soybean and Breaks the Maize-Wheat Paradigm by Engineering a Superior Rhizosphere to Boost Soil and Yield
},
journal = {Advances in Applied Sciences},
volume = {10},
number = {3},
pages = {74-87},
doi = {10.11648/j.aas.20251003.14},
url = {https://doi.org/10.11648/j.aas.20251003.14},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.aas.20251003.14},
abstract = {The intensive maize-wheat double-cropping system underpins food security in China's Huang-Huai-Hai Plain but drives soil degradation through nutrient depletion and biodiversity loss, necessitating sustainable diversification. This study mechanistically compared legacy effects of five preceding summer crops-maize (control), soybean, peanut, pepper, sweet potato-on subsequent winter wheat performance, explicitly quantifying impacts on root architecture, soil enzymatic activity, and yield formation. Results demonstrated peanut’s unparalleled efficacy: it increased seedling-stage wheat shoot biomass by 37-41% and root biomass by 184% versus maize, while expanding root surface area (51%) and volume (54%) through optimized rhizosphere engineering. These morphological advantages persisted through maturity and correlated with significantly enhanced soil functionality-peanut elevated soil organic matter (25-37%), nitrate-N (138-148%), and ammonium-N (71-128%) while reducing C:N ratio. Crucially, peanut residues stimulated microbial metabolism, increasing β-glucosidase activity (governing C cycling) by 33-89% and urease activity (N mineralization) by 40-109%, whereas catalase activity showed context-dependent responses. This accelerated nutrient mineralization translated to agronomic superiority: peanut-wheat rotation yielded 10.5% more grain than maize-wheat, exceeding soybean-wheat by 3.4% despite lower 1000-grain weight, primarily through 26.6% higher ear density. Soybean provided intermediate soil N benefits but weaker root stimulation, while pepper suppressed enzymes and sweet potato inconsistently affected fertility. We conclude that peanut’s unique residue composition-low C:N ratio, abundant labile carbon, and rhizodeposits-primes a self-reinforcing root-microbe-soil loop that enhances nutrient synchrony, breaks maize-wheat yield ceilings, and offers a validated pathway for ecological intensification in cereal systems.},
year = {2025}
}
TY - JOUR T1 - Peanut Precursor Surpasses Soybean and Breaks the Maize-Wheat Paradigm by Engineering a Superior Rhizosphere to Boost Soil and Yield AU - Liu Hecheng AU - Hu Xinru AU - Wang Shancong AU - Reda Mohamed Mahmoud Ahmed AU - Ebtesam Eid Mohamed Abdelhadi AU - Yu Qihang AU - Chen Meiyu AU - Meng Fanzheng AU - Wang Bo AU - Tang Yulou AU - Nasr Mahmoud Abdou AU - Kou Yanling AU - Zhang Rui AU - Shao Ruixin Y1 - 2025/08/08 PY - 2025 N1 - https://doi.org/10.11648/j.aas.20251003.14 DO - 10.11648/j.aas.20251003.14 T2 - Advances in Applied Sciences JF - Advances in Applied Sciences JO - Advances in Applied Sciences SP - 74 EP - 87 PB - Science Publishing Group SN - 2575-1514 UR - https://doi.org/10.11648/j.aas.20251003.14 AB - The intensive maize-wheat double-cropping system underpins food security in China's Huang-Huai-Hai Plain but drives soil degradation through nutrient depletion and biodiversity loss, necessitating sustainable diversification. This study mechanistically compared legacy effects of five preceding summer crops-maize (control), soybean, peanut, pepper, sweet potato-on subsequent winter wheat performance, explicitly quantifying impacts on root architecture, soil enzymatic activity, and yield formation. Results demonstrated peanut’s unparalleled efficacy: it increased seedling-stage wheat shoot biomass by 37-41% and root biomass by 184% versus maize, while expanding root surface area (51%) and volume (54%) through optimized rhizosphere engineering. These morphological advantages persisted through maturity and correlated with significantly enhanced soil functionality-peanut elevated soil organic matter (25-37%), nitrate-N (138-148%), and ammonium-N (71-128%) while reducing C:N ratio. Crucially, peanut residues stimulated microbial metabolism, increasing β-glucosidase activity (governing C cycling) by 33-89% and urease activity (N mineralization) by 40-109%, whereas catalase activity showed context-dependent responses. This accelerated nutrient mineralization translated to agronomic superiority: peanut-wheat rotation yielded 10.5% more grain than maize-wheat, exceeding soybean-wheat by 3.4% despite lower 1000-grain weight, primarily through 26.6% higher ear density. Soybean provided intermediate soil N benefits but weaker root stimulation, while pepper suppressed enzymes and sweet potato inconsistently affected fertility. We conclude that peanut’s unique residue composition-low C:N ratio, abundant labile carbon, and rhizodeposits-primes a self-reinforcing root-microbe-soil loop that enhances nutrient synchrony, breaks maize-wheat yield ceilings, and offers a validated pathway for ecological intensification in cereal systems. VL - 10 IS - 3 ER -
Key Laboratory of High-Efficiency Production of Wheat-Maize Double Cropping, College of Agronomy, Henan Agricultural University, Zhengzhou, China
Key Laboratory of High-Efficiency Production of Wheat-Maize Double Cropping, College of Agronomy, Henan Agricultural University, Zhengzhou, China
School of Modern Horticulture, Henan Vocational College of Agriculture, Zhengzhou, China
Henan Agricultural Technology Extension Central Station, Zhengzhou, China
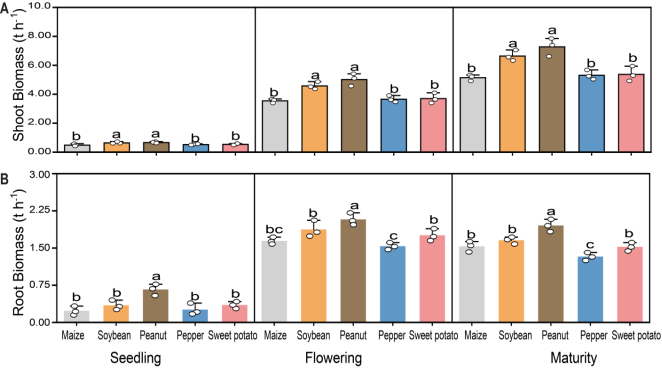
Figure 1. Effects of different preceding crops on (A) shoot and (B) root biomass of winter wheat. Measurements were taken at the seedling, flowering, and maturity stages. Values are means ± standard error (5 plants per replicate x 3 replicates). Different letters above bars indicate significant differences among treatments within a given growth stage according to Fisher's LSD test at P < 0.05.
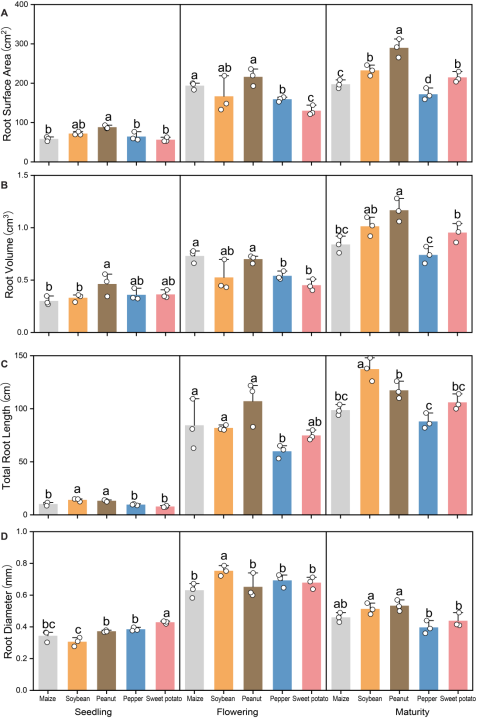
Figure 2. Effects of different preceding crops on winter wheat root morphology: (A) root surface area, (B) root volume, (C) total root length, and (D) average root diameter. Measurements were taken at the seedling, flowering, and maturity stages. Values are means ± standard error (5 plants per replicate x 3 replicates). Different letters above bars indicate significant differences among treatments within a given growth stage at P < 0.05.
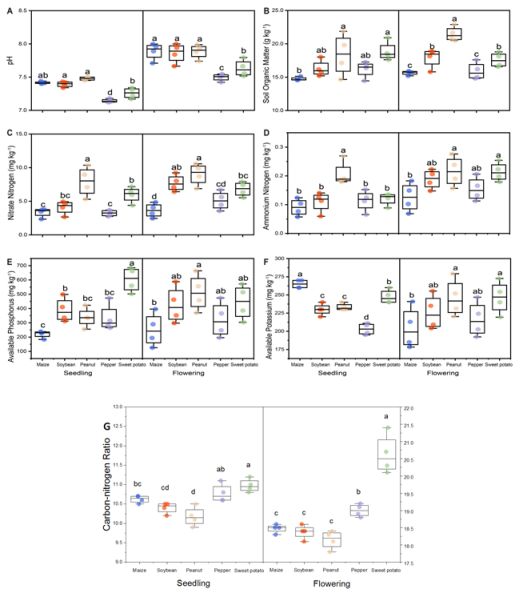
Figure 3. Effects of different preceding crops on soil chemical properties: (A) pH, (B) soil organic matter (SOM), (C) nitrate nitrogen (NO3--N), (D) ammonium nitrogen (NH4+-N), (E) available phosphorus, (F) available potassium, and (G) carbon-nitrogen ratio. Soil samples (0-20 cm) were collected at the winter wheat seedling and flowering stages. Values are means ± standard error (6 sample cores per replicate x 3 replicates). Different letters above bars indicate significant differences among treatments within a given growth stage at P < 0.05.
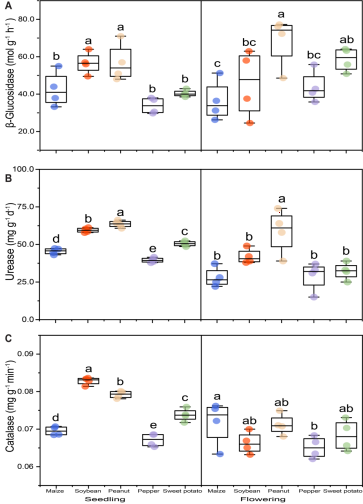
Figure 4. Effects of different preceding crops on soil enzyme activities: (A) β-glucosidase, (B) urease, and (C) catalase. Soil samples (0-20 cm) were collected at the winter wheat seedling and flowering stages. Values are means ± standard error (6 sample cores per replicate x 3 replicates). Different letters above bars indicate significant differences among treatments within a given growth stage at P < 0.05.
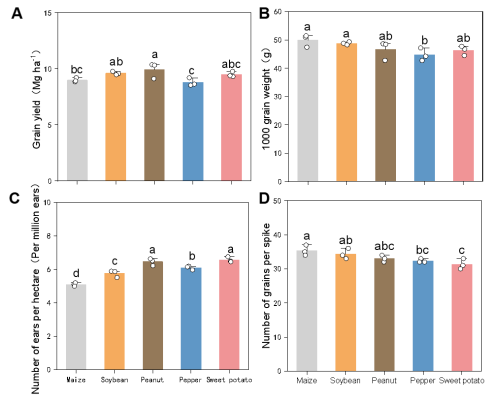
Figure 5. Effects of different preceding crops on winter wheat (A) grain yield, (B) 1000 grain weight, (C) number of ears per hectare and (D) number of grains per spike at harvest. Values are means ± standard error (1 m2 per replicate x 3 replicates). Different letters above bars indicate significant differences among treatments at P < 0.05.
Information

