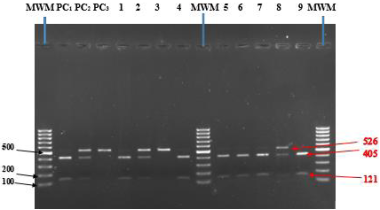
The intrinsic host defense system plays an important role in the restriction of retroviral infection. The objective of the study was to determine the role of Trim5α polymorphism in children living in the North region of Cameroon. A case control study was carried out in the health facilities among HIV-1 infected and uninfected children under 15 years, all born from HIV-1 infected mothers. Blood sample was collected to determine HIV status and genotyping conducted by Polymerase Chain Reaction (PCR) followed by Restriction Fragment Length Polymorphism (RFLP). The Chi-squared test was used to assess the Hardy-Weinberg equilibrium. Overall, 25 HIV-1 infected and 88 uninfected children were recruited. We found that the proportion of GG genotype was lower in uninfected children (85.2%) than in infected ones (92.5%). The proportion of GA genotype was higher in uninfected (12.5%) compared to infected children (8.0%). AA genotype was absent among infected children while the proportion in uninfected children was 2.3%. The frequency of Trim5α-136Q allele in uninfected and infected children was 9.0% and 4.0% respectively. The proportion of mutant homozygotes was elevated in uninfected children (14.8%) than in infected ones (8.0%). Moreover, children carrying mutated phenotype were 2 times less likely to be infected compared to those without it. The mutated phenotype of the Trim5α-136Q gene may be protective against HIV-1 acquisition in children. Further investigation in a follow-up cohort considering other polymorphisms in a large population will help in better appreciation of Trim5α role in HIV-1 acquisition and disease progression in children.
| Published in | Biochemistry and Molecular Biology (Volume 11, Issue 2) |
| DOI | 10.11648/j.bmb.20261102.12 |
| Page(s) | 36-41 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2026. Published by Science Publishing Group |
Genes Variants, HIV-1, MTCT, Restriction Factor, Trim5α
Genes variants | Uninfected children N (%) | Infected children N (%) | P-value | 95%CI | X2 | Total N (%) |
|---|---|---|---|---|---|---|
G/G | 75 (85.2) | 23 (92.0) | 0.58 | -0.22; 0.09 | 0.30 | 98 (86.7) |
G/A | 11 (12.5) | 2 (8.0) | 0.79 | -0.11; 0.20 | 0.07 | 13 (11.5) |
A/A | 2 (2.3) | 0 (0.0) | 0.60 | NA | NA | 2 (1.8) |
total | 88 (100.0) | 25 (100.0) | 113 | |||
Trim5α -136Q frequencies | 9.0 | 4.0 | 8.0 | |||
X2-HWE | 3.46 | 0.04 | 3.39 | |||
P-HWE | 0.06 | 0.83 | 0.07 |
children phenotypes | Uninfected children N (%) | Infected children N (%) | OR | P |
|---|---|---|---|---|
Mutant | 13 (14.8) | 2 (8.0) | 2.0 (0.42-9.50) | 0.29 |
Wild type | 75(85.2) | 23(92.0) | 1 |
AIDS | Acquired Immunodeficiency Syndrome |
DNA | Deoxyribonucleic Acid |
HIV-1 | Human Immunodeficiency Virus Type 1 |
HWE | Hardy-Weinberg Equilibrium |
mRNA | Messenger Ribonucleic Acid |
MTCT | Mother-to-child Transmission |
PCR | Polymerase Chain Reaction |
RFLP | Restriction Fragment Length Polymorphism |
Trim5α | Tripartite Motif Containing 5 Alpha |
| [1] | Spada, S. J., Grigg, M. E., Bouamr, F., Best, S. M., Zhang, P. TRIM5α: A Protean Architect of Viral Recognition and Innate Immunity. Viruses. 2024, 16(7), 997. |
| [2] | Wang, T., Becker, D., Twizerimana, A. P., Luedde, T., Gohlke, H., Münk, C. Cyclophilin ARegulates Tripartite Motif 5 Alpha Restriction of HIV-1. International Journal of Molecular Sciences. 2025, 1-28. |
| [3] | Amanya, S. B., Nyiro, B., Waswa, F., Obura, B., Nakaziba, R., Nabulime, E., Katabazi, A. F., Nabatanzi, R., Bayiyana, A., Mboowa, G., Kayongo, A., Wayengera, M., Sande, O. J. Variations in Trim5α and Cyclophilin A genes among HIV-1 elite controllers and non controllers in Uganda: a laboratory-based cross-sectional study. Retrovirology 17(2020): 1-11. |
| [4] | Ghimire, D., Rai, M., Gaur, R. Novel host restriction factors implicated in HIV-1 replication. J Gen Virol. 2018, 99(4): 435-446. |
| [5] | Malim, M. H., Bieniasz, P. D. HIV Restriction Factors and Mechanisms of Evasion. Cold Spring Harb Perspect Med 2. 2012, 99(4): 435-446. |
| [6] | Blanco-Melo, D., Venkatesh, S., Bieniasz, P. D. Intrinsic cellular defenses against human immunodeficiency viruses. Immunity. 2012, 37(3): 399-411. |
| [7] | Colomer-Lluch, M., Ruiz, A., Moris, A., Prado, J. G. Restriction Factors: From Intrinsic Viral Restriction to Shaping Cellular Immunity Against HIV-1. Front Immunol. 2018, 175-182. |
| [8] | Puvvada, M. P., Patel, S. S. Role of trim5α in the suppression of cross-species transmission and its defence against human immunodeficiency virus. Curr HIV Res. 2013, 11(8): 601-9. |
| [9] | Zhang, X., Kondo, M., Chen, J., Miyoshi, H., Suzuki, H., Ohashi, T., Shida, H. Inhibitory effect of human TRIM5alpha on HIV-1 production. Microbes Infect. 2010, 12(10): 768-77. |
| [10] | Speelmon, E. C., Livingston-Rosanoff, D., Li, S. S., Vu, Q., Bui, J., Geraghty, D. E., Zhao, L. P., McElrath, M. J. Genetic Association of the Antiviral Restriction Factor TRIM5α with Human Immunodeficiency Virus Type 1 Infection. J Virol. 2006, 2463-2471. |
| [11] | Perez-Caballero, D., Hatziioannou, T., Yang, A., Cowan, S., Bieniasz, P. D. Human tripartite motif 5alpha domains responsible for retrovirus restriction activity and specificity. J Virol. 2005, 79(14): 8969-78. |
| [12] | Sokolskaja, E., Berthoux, L., Luban, J. Cyclophilin A and TRIM5alpha independently regulate human immunodeficiency virus type 1 infectivity in human cells. J Virol. 2006, 80(6): 2855-62. |
| [13] | Nakajima, T., Nakayama, E. E., Kaur, G., Terunuma, H., Mimaya, J. I., Ohtani, H., Mehra, N., Shioda, T., Kimura, A. Impact of novel TRIM5alpha variants, Gly110Arg and G176del, on the anti-HIV-1 activity and the susceptibility to HIV-1 infection. AIDS. 2009, 23(16): 2091-100. |
| [14] | Nakayama, E. E., Nakajima, T., Kaur, G., Mimaya J., Terunuma, H., Mehra, N., Kimura, A., Shioda, T. A Naturally Occurring Single Amino Acid Substitution in Human TRIM5a Linker Region Affects Its Anti-HIV Type 1 Activity and Susceptibility to HIV Type 1 Infection. Aids research and human retroviruses, 2013. 29(6): 919-24. |
| [15] | Feng-Liang, L., Yu-Qing, Q., Hong, L., Yi-Qun, K., Xia, T., Guang, C., Nelson Leung, S. T., Yong-Tang, Z. An HIV-1 Resistance Polymorphism in TRIM5α Gene Among Chinese Intravenous Drug Users. JAIDS Journal of Acquired Immune Deficiency Syndromes. 2011, 306-311. |
| [16] | Cloherty, A. P. M., Rader, A. G., Compeer, B., Ribeiro, C. M. S. Human TRIM5α: Autophagy Connects Cell-Intrinsic HIV-1 Restriction and Innate Immune Sensor Functioning. Viruses. 2021, 13(2): 1-32. |
| [17] | Saha, B., Salemi, M., Williams, G. L, Oh, S., Paffett, M. L., Phinney, B., Mandell, M. A. Interactomic analysis reveals a homeostatic role for the HIV restriction factor TRIM5α in mitophagy. Cell Rep. 2022, 39(6): 1-38. |
| [18] | Jimenez-Guardeño, J. M., Apolonia, L., Betancor, G., Malim, M. H. Immunoproteasome activation enables human TRIM5α restriction of HIV-1. Nat Microbiol. 2019, 4(6): 933-940. |
| [19] | Keown, J. R., Black, M. M., Ferron, A., Yap, M., Barnett, M. J., Pearce, F. G., Stoye, J. P., Goldstone, D. C. A helical LC3-interacting region mediates the interaction between the retroviral restriction factor Trim5α and mammalian autophagy-related ATG8 proteins. J Biol Chem. 2018, 293(47): 18378-18386. |
| [20] | Sastri, J., Campbell, E. M. Recent insights into the mechanism and consequences of trim5a retroviral restriction. AIDS Res Hum Retroviruses, 2011, 27(3): 231-8. |
| [21] | van Manen, D., Rits, M. A. N., Beugeling, C., van Dort, K., Schuitemaker, H., Kootstra, N. A. The Effect of Trim5 polymorphisms on the clinical course of HIV-1 infection. PLoS Pathogens. 2008, 4(2): e18. |
| [22] | Price, H., Lacap, P., Tuff, J., Wachihi, C., Kimani, J., Ball, T. B et al. A TRIM5 alpha exon 2 polymorphism is associated with protection from HIV-1 infection in the Pumwani sex worker cohort. AIDS. 2010, 24(12): 1813-21. |
| [23] | Mekue, L. M., Nkenfou, C. N., Dambaya, B., Fotso, I., Nguefack, F., Fainguem, N., Lobe, E. E., Kuiate J. R., Ndjolo., A. Implication of five AIDS related genes in mother-to-child transmission and acquisition of human immunodeficiency virus 1 in Cameroon. Afr j infect dis 13(2018): 1-10. |
| [24] | Djataou, P., Djuidje Ngounoue, M., Nkenfou-Tchinda, C. N., Ngoufack, M. N., Elong, E., Tiga, A., Muluh, C., Kadji Kameni, J., Djaouda, M., Ndjolo, A., Nkenfou, C. N. Low prevalence of HIV in the northern Cameroon: contribution of some AIDS restriction genes and potential implications for gene therapy. Frontiers in genetics. 2024, 01-11. |
| [25] | Dambaya, B., Nkenfou, C. N., Mekue, L., Teto, G., Ngoufack, N., Ambada, G., Flobert, N., Colizzi, V., Alexis, N. TRIM5α 136Q, CCR5 Promoter 59029G And CCR264I Alleles Impact The Progression Of HIV In Children And Adolescents. Appl Clin Genet. 2019, 12: 203-211. |
| [26] | Huthoff, H., Towers, G. J. Restriction of retroviral replication by APOBEC3G/F and TRIM5alpha. Trends Microbiol. 2008, 16(12): 612-9. |
| [27] | Towers, G. J. The control of viral infection by tripartite motif proteins and cyclophilin A. Retrovirology. 2007, 12; 4: 40. |
| [28] | Javanbakht, H., An, P., Gold, B., Petersen, D. C, O'Huigin, C., Nelson, G. W, O'Brien, S. J., Kirk, G. D., Detels, R., Buchbinder, S., Donfield, S., Shulenin, S., Song, B., Perron, M. J, Stremlau, M., Sodroski, J., Dean, M., Winkler, C. Effects of human TRIM5alpha polymorphisms on antiretroviral function and susceptibility to human immunodeficiency virus infection. Virology. 2006, 354(1): 15-27. |
| [29] | Goldschmidt, V., Bleiber, G., May, M., Martinez, R., Ortiz, M., Telenti, A. Swiss HIV Cohort Study. Role of common human TRIM5alpha variants in HIV-1 disease progression. Retrovirology. 2006, 1-17. |
APA Style
Ngoufack, M. N., Nguefack-Tsague, G., Nkenfou, C. N. (2026). Trim5α Genetic Variants and HIV-1 Infection in the North Region of Cameroon. Biochemistry and Molecular Biology, 11(2), 36-41. https://doi.org/10.11648/j.bmb.20261102.12
ACS Style
Ngoufack, M. N.; Nguefack-Tsague, G.; Nkenfou, C. N. Trim5α Genetic Variants and HIV-1 Infection in the North Region of Cameroon. Biochem. Mol. Biol. 2026, 11(2), 36-41. doi: 10.11648/j.bmb.20261102.12
@article{10.11648/j.bmb.20261102.12,
author = {Marie Nicole Ngoufack and Georges Nguefack-Tsague and Celine Nguefeu Nkenfou},
title = {Trim5α Genetic Variants and HIV-1 Infection in the North Region of Cameroon},
journal = {Biochemistry and Molecular Biology},
volume = {11},
number = {2},
pages = {36-41},
doi = {10.11648/j.bmb.20261102.12},
url = {https://doi.org/10.11648/j.bmb.20261102.12},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.bmb.20261102.12},
abstract = {The intrinsic host defense system plays an important role in the restriction of retroviral infection. The objective of the study was to determine the role of Trim5α polymorphism in children living in the North region of Cameroon. A case control study was carried out in the health facilities among HIV-1 infected and uninfected children under 15 years, all born from HIV-1 infected mothers. Blood sample was collected to determine HIV status and genotyping conducted by Polymerase Chain Reaction (PCR) followed by Restriction Fragment Length Polymorphism (RFLP). The Chi-squared test was used to assess the Hardy-Weinberg equilibrium. Overall, 25 HIV-1 infected and 88 uninfected children were recruited. We found that the proportion of GG genotype was lower in uninfected children (85.2%) than in infected ones (92.5%). The proportion of GA genotype was higher in uninfected (12.5%) compared to infected children (8.0%). AA genotype was absent among infected children while the proportion in uninfected children was 2.3%. The frequency of Trim5α-136Q allele in uninfected and infected children was 9.0% and 4.0% respectively. The proportion of mutant homozygotes was elevated in uninfected children (14.8%) than in infected ones (8.0%). Moreover, children carrying mutated phenotype were 2 times less likely to be infected compared to those without it. The mutated phenotype of the Trim5α-136Q gene may be protective against HIV-1 acquisition in children. Further investigation in a follow-up cohort considering other polymorphisms in a large population will help in better appreciation of Trim5α role in HIV-1 acquisition and disease progression in children.},
year = {2026}
}
TY - JOUR T1 - Trim5α Genetic Variants and HIV-1 Infection in the North Region of Cameroon AU - Marie Nicole Ngoufack AU - Georges Nguefack-Tsague AU - Celine Nguefeu Nkenfou Y1 - 2026/06/27 PY - 2026 N1 - https://doi.org/10.11648/j.bmb.20261102.12 DO - 10.11648/j.bmb.20261102.12 T2 - Biochemistry and Molecular Biology JF - Biochemistry and Molecular Biology JO - Biochemistry and Molecular Biology SP - 36 EP - 41 PB - Science Publishing Group SN - 2575-5048 UR - https://doi.org/10.11648/j.bmb.20261102.12 AB - The intrinsic host defense system plays an important role in the restriction of retroviral infection. The objective of the study was to determine the role of Trim5α polymorphism in children living in the North region of Cameroon. A case control study was carried out in the health facilities among HIV-1 infected and uninfected children under 15 years, all born from HIV-1 infected mothers. Blood sample was collected to determine HIV status and genotyping conducted by Polymerase Chain Reaction (PCR) followed by Restriction Fragment Length Polymorphism (RFLP). The Chi-squared test was used to assess the Hardy-Weinberg equilibrium. Overall, 25 HIV-1 infected and 88 uninfected children were recruited. We found that the proportion of GG genotype was lower in uninfected children (85.2%) than in infected ones (92.5%). The proportion of GA genotype was higher in uninfected (12.5%) compared to infected children (8.0%). AA genotype was absent among infected children while the proportion in uninfected children was 2.3%. The frequency of Trim5α-136Q allele in uninfected and infected children was 9.0% and 4.0% respectively. The proportion of mutant homozygotes was elevated in uninfected children (14.8%) than in infected ones (8.0%). Moreover, children carrying mutated phenotype were 2 times less likely to be infected compared to those without it. The mutated phenotype of the Trim5α-136Q gene may be protective against HIV-1 acquisition in children. Further investigation in a follow-up cohort considering other polymorphisms in a large population will help in better appreciation of Trim5α role in HIV-1 acquisition and disease progression in children. VL - 11 IS - 2 ER -
Department of Chemistry-Biology, University of Bertoua, Bertoua, Cameroon
Information