This study is geared towards investigating stability analysis, dispersion relation and pattern formation potential of damage in the body of living organisms using 3-coupled nonlinear system of partial differential equations (PDEs). Obviously, this study proved that there are possibilities of asymptomatic stage of the damage with the first equilibrium point and the second is symptomatic stage of damaged cells at equilibrium which is usually and physically experienced in diabetic patients. The second equilibrium point predicts initiation of damaged cells, progression or inhibitions for healing. The stability analysis revealed steady state solutions with the exudation surface activated by the stress, temperature and viscous terms causing increased growth rate of damaged cells in the body of the organism. The solutions of dispersion relations is an archetype of spatial heterogeneous and homogeneous solutions of the linear biological systems with unbalanced and exponential growth term linearly (linear growth rate of damaged cells and its wave vector k) with respect to time. The study identified that the quadratic nature of Ψ(k2) would lead the system to bounded finite amplitude. The study explored into complex dispersion relations and ascertained some special cases where one or more parameters affected damaged cells mechanically at equilibrium with bifurcation surface between spatial homogeneity and heterogeneity. Here, the bifurcation parameter τ increases monotonically beyond the critical value and renders B(k2) zero and the uniform steady state bifurcates to a spatial unstable state. This study showed pattern formation potentials are quadratic and bounded domain of the injured cells, spatial pattern of random and viscous initial damaged cell densities and patterning process of no uniform damaged cell density due cell traction is bigger than the critical value of cell traction which is the resistance of the extracellular matrix and spatial non-homogeneities activates which grows out of bounded region for some k. The study used partial Differential equation and determinant methods to proffer the solutions. Among others, this study recommends that the affected individuals should explore the use of antioxidants as to combat with the out growing inbuilt stress called oxidative stress which is activator of damaged cells densities in the body of the organisms.
| Published in | International Journal of Applied Mathematics and Theoretical Physics (Volume 12, Issue 1) |
| DOI | 10.11648/j.ijamtp.20261201.15 |
| Page(s) | 44-54 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2026. Published by Science Publishing Group |
Biological Tissues, Body, Damage, Dispersion Relation, Living Organisms, Pattern Formation, Stability Analysis
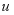 small, upon substituting into the nonlinear system of equation and keeping the linear terms in, and , , and is a measure of cell –cell contact activation of cancer cells and also nonnegative. Setting the solutions the linearized equations to be
small, upon substituting into the nonlinear system of equation and keeping the linear terms in, and , , and is a measure of cell –cell contact activation of cancer cells and also nonnegative. Setting the solutions the linearized equations to be ODE | Ordinary Differential Equations |
ECM | Extra-Cellular Matrix |
PDE | Partial Differential Equation |
| [1] | Alonso S, Radszuweit M, Engel H. & Bär M. (2017). Mechanochemical pattern formation in simple models of active viscoelastic fluids and solids. J Phys D Appl Phys. 50(43): 434004. |
| [2] | Brinkmann, F., Mercker, M., Richter, T. & Marciniak-Czochra, A. (2018). Post-turing tissue pattern formation: advent of mechanochemistry. PLoS Comput Biol. 14(7): e1006259. |
| [3] | Byrne, H. & Preziosi, L.(2003). Modelling solid tumour growth using the theory of mixtures. Math Med Biol J IMA. 20(4): 341–366. |
| [4] | Chandler C, Liu T, Buckanovich R, Coffman L. (2019). The double edge sword of fibrosis in cancer. Trans Res. 209: 55–67. |
| [5] | Chaplain MAJ, V. C.; Gerisch, A. & Lorenzi, T. (2021). Mechanical Models of Pattern and Form in Biological Tissues: The Role of Stress-Strain Constitutive Equations. Bull Math Biol. 83(7): 80. |
| [6] | Cross M. & Greenside H. (2009). Pattern formation and dynamics in nonequilibrium systems. Cambridge: Cambridge University Press. |
| [7] | Ebihara T, Venkatesan N, Tanaka R, Ludwig MS. (2000). Changes in extracellular matrix and tissue viscoelasticity in bleomycin-induced lung fibrosis: Temporal aspects. Am J Respirat Crit Care Med. 162(4): 1569–1576. |
| [8] | Gilmore SJ, Vaughan BL, Jr, Madzvamuse A. & Maini, PK. (2012). A mechanochemical model of striae distensae. Math Biosci. 240(2): 141–147. |
| [9] | Haghighi-Yazdi M. & Lee-Sullivan P (2011) Modeling linear viscoelasticity in glassy polymers using standard rheological models. |
| [10] | Hylan, KM. (2014.). Tumor suppressor genes and oncogenes: Genes that prevent and cause cancer. |
| [11] | Javierre E, Moreo P, Doblaré M.& García-Aznar JM. (2009). Numerical modeling of a mechano–chemical theory for wound contraction analysis. Int J Solids Struct. 46(20): 3597–3606. |
| [12] | Jezierska–Drutel, A., Rosenzweig, S. A. & Neumann, C. A. (2013). Role of Oxidative Stress and the Microenvironment in Breast Cancer Development and Progression. Advances in Cancer Research, Vol 119, Elsevier Inc. |
| [13] | Khalilgharibi N. & Mao Y. (2021). To form and function: on the role of basement membrane mechanics in tissue development, homeostasis and disease. Open Biol. 11(2): 200360. |
| [14] | Konieczny, L., Roterman-Konieczna, I., Spólnik, P. (2023). The Structure and Function of Living Organisms. In: Systems Biology. Springer, Cham. |
| [15] | Maini, P. K. & Woolley, T. E. (2019). The Turing model for biological pattern formation. In: The dynamics of biological systems. SpringeR, 189–204. |
| [16] | Maini PK, Woolley TE, Baker RE, Gaffney EA, Lee SS. (2012). Turing’s model for biological pattern formation and the robustness problem. Interface Focus. 2(4): 487–496. |
| [17] | Moreo P, Gaffney EA, Garcia-Aznar JM, Doblaré M. (2010). On the modelling of biological patterns with mechanochemical models: insights from analysis and computation. Bull Math Biol. 72(2): 400–431. |
| [18] | Marray, J. D. (2003). Mathematical Biology II: Spatial Models and Biomedical Applications. 3rd ed. 18(1), Springer New York. |
| [19] | Petrolli V, Le Goff M, Tadrous M, Martens K, Allier C, Mandula O, Hervé L, Henkes S, Sknepnek R. & Boudou T, et al. (2019). Confinement-induced transition between wavelike collective cell migration modes. Phys Rev Lett. 122(16): 168101. |
| [20] | Rodrigo, S. G., Hastings, P. J. and Rosenberg, S. M.(2012). Mutation as a Stress Response and the Regulation of Evolvability. HHS Public Access. |
| [21] | Scianna M, Bell CG, Preziosi L. (2013). A review of mathematical models for the formation of vascular networks. J Theor Biol. 333: 174–209. |
| [22] | Serra-Picamal X, Conte V, Vincent R, Anon E, Tambe DT, Bazellieres E, Butler JP, Fredberg JJ, Trepat X. (2012). Mechanical waves during tissue expansion. Nat Phys. 8(8): 628–634. |
| [23] | Shain, A. H. & Pollack, J. R.(2013). The spectrum of SWI/SNF mutations, ubiquitous in Human Cancer. PLoS ONE. 8(1): e55119. |
| [24] | Tlili S, Gauquelin E, Li B, Cardoso O, Ladoux B, Delanoë-Ayari H. & Graner F. (2018). Collective cell migration without proliferation: density determines cell velocity and wave velocity. R Soc Open Sci. 5(5): 172421. |
| [25] | Urdy, S. (2012). On the evolution of morphogenetic models: mechano-chemical interactions and an integrated view of cell differentiation, growth, pattern formation and morphogenesis. Biol Rev. 87(4): 786–803. |
| [26] | Van Helvert, S. Storm, C. & Friedl, P. (2018). Mechanoreciprocity in cell migration. Nat Cell Biol. 20(1): 8–20. |
APA Style
Nkuturum, C., Ban, T. S., Egharevba, A. J., George, L. E. (2026). Stability Analysis, Dispersion Relation and Pattern Formation of Damaged Cells on Biological Tissues in the Body of Living Organisms. International Journal of Applied Mathematics and Theoretical Physics, 12(1), 44-54. https://doi.org/10.11648/j.ijamtp.20261201.15
ACS Style
Nkuturum, C.; Ban, T. S.; Egharevba, A. J.; George, L. E. Stability Analysis, Dispersion Relation and Pattern Formation of Damaged Cells on Biological Tissues in the Body of Living Organisms. Int. J. Appl. Math. Theor. Phys. 2026, 12(1), 44-54. doi: 10.11648/j.ijamtp.20261201.15
@article{10.11648/j.ijamtp.20261201.15,
author = {Christiana Nkuturum and Tombari Stephen Ban and Aimuamwosa Julia Egharevba and Lauretta Emugha George},
title = {Stability Analysis, Dispersion Relation and Pattern Formation of Damaged Cells on Biological Tissues in the Body of Living Organisms},
journal = {International Journal of Applied Mathematics and Theoretical Physics},
volume = {12},
number = {1},
pages = {44-54},
doi = {10.11648/j.ijamtp.20261201.15},
url = {https://doi.org/10.11648/j.ijamtp.20261201.15},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.ijamtp.20261201.15},
abstract = {This study is geared towards investigating stability analysis, dispersion relation and pattern formation potential of damage in the body of living organisms using 3-coupled nonlinear system of partial differential equations (PDEs). Obviously, this study proved that there are possibilities of asymptomatic stage of the damage with the first equilibrium point and the second is symptomatic stage of damaged cells at equilibrium which is usually and physically experienced in diabetic patients. The second equilibrium point predicts initiation of damaged cells, progression or inhibitions for healing. The stability analysis revealed steady state solutions with the exudation surface activated by the stress, temperature and viscous terms causing increased growth rate of damaged cells in the body of the organism. The solutions of dispersion relations is an archetype of spatial heterogeneous and homogeneous solutions of the linear biological systems with unbalanced and exponential growth term linearly (linear growth rate of damaged cells and its wave vector k) with respect to time. The study identified that the quadratic nature of Ψ(k2) would lead the system to bounded finite amplitude. The study explored into complex dispersion relations and ascertained some special cases where one or more parameters affected damaged cells mechanically at equilibrium with bifurcation surface between spatial homogeneity and heterogeneity. Here, the bifurcation parameter τ increases monotonically beyond the critical value and renders B(k2) zero and the uniform steady state bifurcates to a spatial unstable state. This study showed pattern formation potentials are quadratic and bounded domain of the injured cells, spatial pattern of random and viscous initial damaged cell densities and patterning process of no uniform damaged cell density due cell traction is bigger than the critical value of cell traction which is the resistance of the extracellular matrix and spatial non-homogeneities activates which grows out of bounded region for some k. The study used partial Differential equation and determinant methods to proffer the solutions. Among others, this study recommends that the affected individuals should explore the use of antioxidants as to combat with the out growing inbuilt stress called oxidative stress which is activator of damaged cells densities in the body of the organisms.},
year = {2026}
}
TY - JOUR T1 - Stability Analysis, Dispersion Relation and Pattern Formation of Damaged Cells on Biological Tissues in the Body of Living Organisms AU - Christiana Nkuturum AU - Tombari Stephen Ban AU - Aimuamwosa Julia Egharevba AU - Lauretta Emugha George Y1 - 2026/03/14 PY - 2026 N1 - https://doi.org/10.11648/j.ijamtp.20261201.15 DO - 10.11648/j.ijamtp.20261201.15 T2 - International Journal of Applied Mathematics and Theoretical Physics JF - International Journal of Applied Mathematics and Theoretical Physics JO - International Journal of Applied Mathematics and Theoretical Physics SP - 44 EP - 54 PB - Science Publishing Group SN - 2575-5927 UR - https://doi.org/10.11648/j.ijamtp.20261201.15 AB - This study is geared towards investigating stability analysis, dispersion relation and pattern formation potential of damage in the body of living organisms using 3-coupled nonlinear system of partial differential equations (PDEs). Obviously, this study proved that there are possibilities of asymptomatic stage of the damage with the first equilibrium point and the second is symptomatic stage of damaged cells at equilibrium which is usually and physically experienced in diabetic patients. The second equilibrium point predicts initiation of damaged cells, progression or inhibitions for healing. The stability analysis revealed steady state solutions with the exudation surface activated by the stress, temperature and viscous terms causing increased growth rate of damaged cells in the body of the organism. The solutions of dispersion relations is an archetype of spatial heterogeneous and homogeneous solutions of the linear biological systems with unbalanced and exponential growth term linearly (linear growth rate of damaged cells and its wave vector k) with respect to time. The study identified that the quadratic nature of Ψ(k2) would lead the system to bounded finite amplitude. The study explored into complex dispersion relations and ascertained some special cases where one or more parameters affected damaged cells mechanically at equilibrium with bifurcation surface between spatial homogeneity and heterogeneity. Here, the bifurcation parameter τ increases monotonically beyond the critical value and renders B(k2) zero and the uniform steady state bifurcates to a spatial unstable state. This study showed pattern formation potentials are quadratic and bounded domain of the injured cells, spatial pattern of random and viscous initial damaged cell densities and patterning process of no uniform damaged cell density due cell traction is bigger than the critical value of cell traction which is the resistance of the extracellular matrix and spatial non-homogeneities activates which grows out of bounded region for some k. The study used partial Differential equation and determinant methods to proffer the solutions. Among others, this study recommends that the affected individuals should explore the use of antioxidants as to combat with the out growing inbuilt stress called oxidative stress which is activator of damaged cells densities in the body of the organisms. VL - 12 IS - 1 ER -
Department of Mathematics, Rivers State University Nkpolu-Oroworukwo, Port Harcourt, Nigeria
Department of Mathematics, Rivers State University Nkpolu-Oroworukwo, Port Harcourt, Nigeria
Department of Mathematics and Statistics, University of Port Harcourt, Port Harcourt, Nigeria
Department of Mathematics and Statistics, Ignatius Ajuru University of Education, Port Harcourt, Nigeria
Information

