Maintenance of genomic integrity is essential for all organism that help to maintain the number of chromosomes during cell division. In yeast and other organisms, the diploidization of genome serves as a key survival strategy under changing environmental conditions. Fission yeast ATR homolog Rad3 and TOR complex protein Tor2 are phosphatidylinositol 3-kinase-related protein kinases (PIKKs) and are involved in various functions. Rad3 is a key regulator of DNA damage signaling pathway and is involved in the maintenance of genomic integrity. The TOR complex, TORC1 and TORC2, contain Tor2 and Tor1 protein kinase respectively as their major component are involved in regulating eukaryotic cell growth, metabolism, and survival, in response to nutrient, energy, and environmental cues. In this study, we observed the suppression of temperature sensitive phenotype of a tor2-287 mutation in rad3Δ background. The presence of azygotic asci and tetrad dissection analysis revealed the diploid nature of rad3Δ tor2-287 double mutant cells, which was further confirmed by flow cytometry (FACS) analysis. In addition, the tor2-287/tor2-287 homozygous and tor-287/tor2+ heterozygous diploid exhibit temperature sensitive and resistant phenotypes, respectively. The tetrad dissection analysis revealed that a wild type copy of the tor2+ and rad3+ genes was present in tor2-287 rad3Δ strain due to the diploid nature of these cells, thereby suppressing the temperature sensitive phenotype. Overall, our results demonstrate that the absence of rad3 suppresses the temperature sensitivity of tor2-287 mutant by maintaining the diploid nature of cells.
| Published in | International Journal of Genetics and Genomics (Volume 14, Issue 1) |
| DOI | 10.11648/j.ijgg.20261401.14 |
| Page(s) | 39-44 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2026. Published by Science Publishing Group |
Rad3, Tor2, Ploidy, S. pombe
Strain | Genotype | Source |
|---|---|---|
SP3 | h+ leu1-32 | Lab stock |
SH1695 | h+ leu1-32 ura4D18 rad3:: ura4 | This study, derived from FY14154 (NBRP) |
SH1692 | h- ura4D18 tor2-287:: hygR | This study, derived from FY40338 (NBRP) |
SH1701 | h+ leu1-32 ura4D18 tor2-287:: hygR rad3:: ura4 (exist as diploid) | This study |
SH1836 | h leu1-32 ura4D18 tor2-287:: hygR rad3:: ura4 (exist as diploid) | This study |
SH2094 | h+/h- leu1-32/leu1+ tor2-287:: hygR/tor2-287:: hygR ade6-216/ade6-210 | This study |
SH2096 | h+/h- leu1-32/leu1+ tor2-287:: hygR/tor2+ ade6-216/ade6-210 | This study |
SH1658 | h+/h- leu1-32/leu1-32 ade6-216/ade6-210 | Lab stock |
S.No. | Genotype | Total no. of cells | Azygotic asci | Zygotic asci | % azygotic asci |
|---|---|---|---|---|---|
1 | Wild type | 251 | 0 | 1 | 0.4% |
2 | rad3::ura4 | 222 | 0 | 1 | 0.45% |
3 | tor2-287::hygR | 212 | 0 | 0 | 0 |
4 | tor2-287::hygR rad3::ura4 | 295 | 56 | 12 | 19% |
5 | Wild type diploid | 185 | 44 | 9 | 23.8% |
FACS | Flow Cytometry |
YEA | Yeast Extract Agar |
EMM | Edinburgh Minimal Media |
YES | Yeast Extract with Suppliment |
DNA | Deoxyribonucleic Acid |
DIC | Differential Interference Contrast Microscopy |
PIKK | Phosphatidylinositol 3-Kinase-Related Protein Kinase |
| [1] | Storchova, Z., Pellman, D., From polyploidy to aneuploidy, genome instability and cancer, Nat Rev Mol Cell Biol, 2004, vol. 5, pp. 45–54. |
| [2] | Selmecki, A., Forche, A., BermanJ., Aneuploidy, and, isochromosome formation, in drug-resistant Candida albicans, Science, 2006, vol. 313, pp. 367–370. |
| [3] | Selmecki, A. M., Maruvka, Y. E., Richmond, P. A., Guillet, M., Shoresh, N., Sorenson, A. L., De, S., Kishony, R., Michor, F., Dowell, R. and Pellman, D., Polyploidy can drive rapid adaptation in yeast, Nature, 2015, vol. 519 (7543), pp. 349-352. |
| [4] | Bentley, N. J., Holtzman, D. A., Flaggs, G., Keegan, K. S., DeMaggio, A., Ford, J. C., Hoekstra, M., Carr, A. M., The Schizosaccharomyces pombe Rad3 Checkpoint Gene, EMBO J, 1996, vol. 15, pp. 6641–6651. |
| [5] | Awasthi, P., Foiani, M., Kumar, A., ATM and ATR signaling at a glance, J Cell Sci, 2015, vol. 128(23), pp. 4255-4262. |
| [6] | Ahamad, N., Sharma, T., Khan, S., Siddiqi, M. I., Ahmed, S., Phosphorylation of Wat1, human Lst8 homolog is critical for the regulation of TORC2 –Gad8 dependent pathway in fission yeast Schizosaccharomyces pombe. European Journal of Cell Biology, 2018, 97(4), pp. 300–307. |
| [7] | Hayashi, T., Hatanaka, M., Nagao, K., Nakaseko, Y., Kanoh, J., Kokubu, A., Ebe, M., Yanagida, M., Rapamycin sensitivity of the Schizosaccharomyces pombe tor2 mutant and organization of two highly phosphorylated TOR complexes by specific and common subunits, Genes Cells, 2007, vol. 12(12), pp. 1357–1370. |
| [8] | Tatebe, H., Shiozaki, K., Evolutionary conservation of the components in the tor signaling pathways. Biomolecules, 2017, 7(4), pp. 1–17. |
| [9] | Morozumi, Y., Shiozaki, K., Conserved and divergent mechanisms that control TORC1 in yeasts and mammals, Genes, 2021, vol. 12(1), pp. 88. |
| [10] | Alvarez, B., Moreno, S., Fission yeast Tor2 promotes cell growth and represses cell differentiation, J Cell Sci, 2006, vol. 119(21), pp. 4475–4485. |
| [11] | Panigrahi, L., Anjum, S., Ahmed, S., Critical role of Wat1/Pop3 in regulating the TORC1 signalling pathway in fission yeast S. pombe, Fungal Genetics and Biology, 2023 vol. 164, pp. 103764. |
| [12] | Anjum, S., Srivastava, S., Panigrahi, L., Ansari, U. A., Trivedi, A. K., Ahmed, S., TORC1 mediated regulation of mitochondrial integrity and calcium ion homeostasis by Wat1/mLst8 in S. pombe. International Journal of Biological Macromolecules, 2023, 253, p. 126907. |
| [13] | Ahamad, N., Verma, S. K., Ahmed, S., Activation of checkpoint kinase Chk1 by reactive oxygen species resulting from disruption of wat1/pop3 in Schizosaccharomyces pombe. Genetics, 2016, 204(4), pp. 1397-1406. |
| [14] | Forsburg, S. L., Rhind, N., Basic methods for fission yeast. Yeast, 2006, 23(3), pp. 173-183. |
| [15] | Khan, S., Ahmed, S., Role of swi7H4 mutant allele of DNA polymerase α in the DNA damage checkpoint response, PLoS One, 2015, vol. 10(3), e0124063. |
| [16] | Klar, A. J., A genetic mechanism implicates chromosome 11 in schizophrenia and bipolar diseases, Genetics, 2004, vol. 167(4), pp. 1833-40. |
| [17] | Dalgaard, J. Z., Klar, A. J., Does S. pombe exploit the intrinsic asymmetry of DNA synthesis to imprint daughter cells for mating-type switching? Trends in Genetics, 2001, vol. 17(3), pp. 153-157. |
| [18] | Harari, Y., Ram, Y., Kupiec, M., Frequent ploidy changes in growing yeast cultures, Curr Genetics, 2018, vol. 64, pp. 1001-1004. |
| [19] | Chen, J., Xiong, Z., Miller, D. E., Yu, Z., McCroskey, S., Bradford, W. D., Cavanaugh, A. M., Jaspersen, S. L., The role of gene dosage in budding yeast centrosome scaling and spontaneous diploidization, PLoS Genet, 2020, vol. 16(12), e1008911. |
| [20] | Stepchenkova, E. I., Shiriaeva, A. A., Pavlov, Y. I., Deletion of the DEF1 gene does not confer UV-immutability but frequently leads to self-diploidization in yeast Saccharomyces cerevisiae, DNA repair, 2018 vol. 70, pp. 49-54. |
| [21] | Tung, S., Bakerlee, C. W., Phillips, A. M., Nguyen, Ba. A. N., Desai, M. M., The genetic basis of differential autodiploidization in evolving yeast populations. G3, 2021, vol. 11(8), jkab192. |
APA Style
Deoralia, M., Panigrahi, L., Ahmed, S. (2026). The Loss of Rad3 Induces the Diploidization of Tor2-287 Mutant to Promote the Cell Survival. International Journal of Genetics and Genomics, 14(1), 39-44. https://doi.org/10.11648/j.ijgg.20261401.14
ACS Style
Deoralia, M.; Panigrahi, L.; Ahmed, S. The Loss of Rad3 Induces the Diploidization of Tor2-287 Mutant to Promote the Cell Survival. Int. J. Genet. Genomics 2026, 14(1), 39-44. doi: 10.11648/j.ijgg.20261401.14
@article{10.11648/j.ijgg.20261401.14,
author = {Monika Deoralia and Lalita Panigrahi and Shakil Ahmed},
title = {The Loss of Rad3 Induces the Diploidization of Tor2-287 Mutant to Promote the Cell Survival},
journal = {International Journal of Genetics and Genomics},
volume = {14},
number = {1},
pages = {39-44},
doi = {10.11648/j.ijgg.20261401.14},
url = {https://doi.org/10.11648/j.ijgg.20261401.14},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.ijgg.20261401.14},
abstract = {Maintenance of genomic integrity is essential for all organism that help to maintain the number of chromosomes during cell division. In yeast and other organisms, the diploidization of genome serves as a key survival strategy under changing environmental conditions. Fission yeast ATR homolog Rad3 and TOR complex protein Tor2 are phosphatidylinositol 3-kinase-related protein kinases (PIKKs) and are involved in various functions. Rad3 is a key regulator of DNA damage signaling pathway and is involved in the maintenance of genomic integrity. The TOR complex, TORC1 and TORC2, contain Tor2 and Tor1 protein kinase respectively as their major component are involved in regulating eukaryotic cell growth, metabolism, and survival, in response to nutrient, energy, and environmental cues. In this study, we observed the suppression of temperature sensitive phenotype of a tor2-287 mutation in rad3Δ background. The presence of azygotic asci and tetrad dissection analysis revealed the diploid nature of rad3Δ tor2-287 double mutant cells, which was further confirmed by flow cytometry (FACS) analysis. In addition, the tor2-287/tor2-287 homozygous and tor-287/tor2+ heterozygous diploid exhibit temperature sensitive and resistant phenotypes, respectively. The tetrad dissection analysis revealed that a wild type copy of the tor2+ and rad3+ genes was present in tor2-287 rad3Δ strain due to the diploid nature of these cells, thereby suppressing the temperature sensitive phenotype. Overall, our results demonstrate that the absence of rad3 suppresses the temperature sensitivity of tor2-287 mutant by maintaining the diploid nature of cells.},
year = {2026}
}
TY - JOUR T1 - The Loss of Rad3 Induces the Diploidization of Tor2-287 Mutant to Promote the Cell Survival AU - Monika Deoralia AU - Lalita Panigrahi AU - Shakil Ahmed Y1 - 2026/03/16 PY - 2026 N1 - https://doi.org/10.11648/j.ijgg.20261401.14 DO - 10.11648/j.ijgg.20261401.14 T2 - International Journal of Genetics and Genomics JF - International Journal of Genetics and Genomics JO - International Journal of Genetics and Genomics SP - 39 EP - 44 PB - Science Publishing Group SN - 2376-7359 UR - https://doi.org/10.11648/j.ijgg.20261401.14 AB - Maintenance of genomic integrity is essential for all organism that help to maintain the number of chromosomes during cell division. In yeast and other organisms, the diploidization of genome serves as a key survival strategy under changing environmental conditions. Fission yeast ATR homolog Rad3 and TOR complex protein Tor2 are phosphatidylinositol 3-kinase-related protein kinases (PIKKs) and are involved in various functions. Rad3 is a key regulator of DNA damage signaling pathway and is involved in the maintenance of genomic integrity. The TOR complex, TORC1 and TORC2, contain Tor2 and Tor1 protein kinase respectively as their major component are involved in regulating eukaryotic cell growth, metabolism, and survival, in response to nutrient, energy, and environmental cues. In this study, we observed the suppression of temperature sensitive phenotype of a tor2-287 mutation in rad3Δ background. The presence of azygotic asci and tetrad dissection analysis revealed the diploid nature of rad3Δ tor2-287 double mutant cells, which was further confirmed by flow cytometry (FACS) analysis. In addition, the tor2-287/tor2-287 homozygous and tor-287/tor2+ heterozygous diploid exhibit temperature sensitive and resistant phenotypes, respectively. The tetrad dissection analysis revealed that a wild type copy of the tor2+ and rad3+ genes was present in tor2-287 rad3Δ strain due to the diploid nature of these cells, thereby suppressing the temperature sensitive phenotype. Overall, our results demonstrate that the absence of rad3 suppresses the temperature sensitivity of tor2-287 mutant by maintaining the diploid nature of cells. VL - 14 IS - 1 ER -
Biochemistry and Structural Biology Division, CSIR- Central Drug Research Institute, Lucknow, India;Academy of Scientific and Innovative Research (AcSIR), Ghaziabad, India
Biochemistry and Structural Biology Division, CSIR- Central Drug Research Institute, Lucknow, India
Biochemistry and Structural Biology Division, CSIR- Central Drug Research Institute, Lucknow, India;Academy of Scientific and Innovative Research (AcSIR), Ghaziabad, India
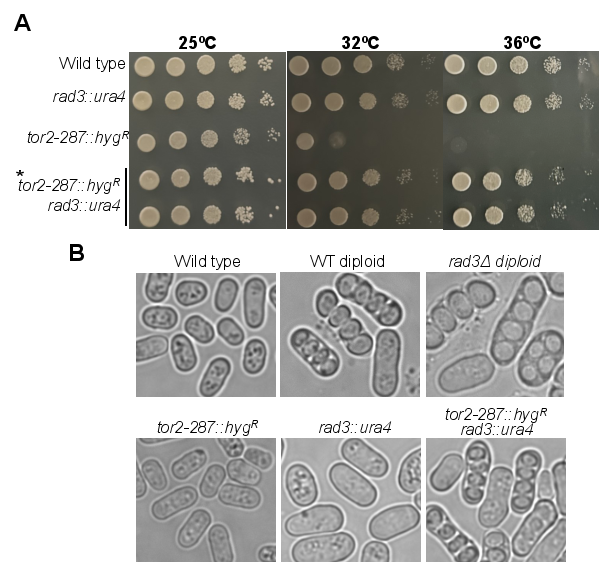
Figure 1. The double mutation of rad3Δ tor2-287 suppresses temperature sensitive phenotype with azygotic asci. (A) Absence of rad3 suppresses the temperature sensitivity of tor2-287 mutant. Indicated strains were grown at 25°C till mid log phase, serially diluted, and spotted on rich media (YES) plates. Plates were allowed to grow at indicated temperature for 3-4 days before taking a photograph. The asterisk denotes a probable diploid nature of cells. The results shown here are from one set of experiment out of three independent experiments (B) Indicated strains were grown on an EMM agar plate, and a DIC image of cells containing spores was monitored under a microscope. For each strain, more than 200 cells were monitored for the calculation of percent azygotic asci.

Figure 2. The double mutant of tor2-287 rad3Δ cells are diploid in nature and exhibits 2:2 segregation. The asci from rad3Δ tor2-287 double mutant were dissected on a YES plate, and replica plated on the indicated plates. Parental strains were also patched and incubated at 25oC or 36oC for 3-4 days before taking a photograph. The results shown here are from one set of dissection out of two independent experiments as described in Methods.
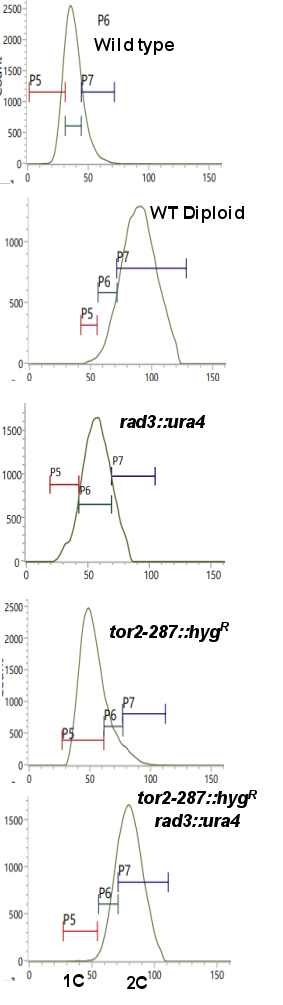
Figure 3. FACS analysis of cells. Indicated strains were grown at 25°C, stained with propidium iodide, and FACS analysis was performed using BD FACS Calibre for the analysis of DNA content. Multiple independent experiments were performed, and the results shown here are from one set of experiments.
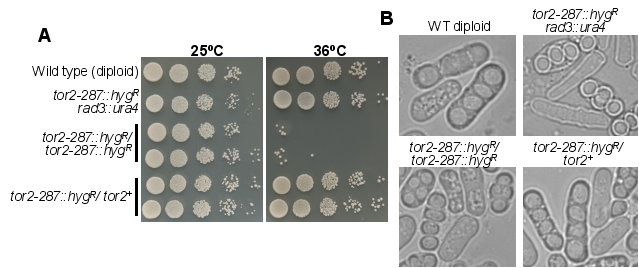
Figure 4. The phenotypic analysis of homozygous and heterozygous diploid strains. (A) The homozygous and heterozygous diploid strains were constructed by genetic crosses as described in Methods, and cells were grown at 25°C till mid log phase, serially diluted, and spotted on rich media (YES) plates. Plates were allowed to grow at the indicated temperature for 3-4 days before taking a photograph. (B) Indicated strains were grown on an EMM agar plate, and a DIC image of cells containing spores was monitored under a microscope.
Information

