TLS/FUS is RNA-binding protein having multiple functions of regulations of genes, homeostasis, and cellular growth. Recent studies show that TLS is involved in phase separation and occasionally forms precipitation related to neurodegenerative diseases like amyotrophic lateral sclerosis (ALS). RNA has been reported to suppress phase separation, droplet formation, and concomitant precipitation of TLS, suggesting that RNA is a possible candidate for ALS drug discovery. Our experiments demonstrated that a long noncoding RNA, promoter-associated noncoding RNA (pncRNA-D), specifically binds TLS and represses its phase separation and precipitation. To obtain competent drug seeds, it is essential to reveal mechanism of action of lncRNAs with specificity to TLS and inhibitory activity on phase separation and related precipitation. For this purpose, several lncRNAs (lncRNAs 1 to 6) were selected upon assays with GST-TLS binding and inhibition on the precipitation. With criteria of binding specificity for TLS, lncRNA3 has been selected for further analysis for RNA-binding ability. Initially, RNA-binding region at TLS amino acid sequence was identified from four fragments of TLS. RNA binding assay with biotinylated lncRNA3 precipitated with avidin magnetic beads indicated clearly that TLS binds the fragment 4 (373-526 aa), C-terminus end of TLS. Then, dissecting fragment 4 presents four regions, RGG2, zinc finger, RGG3, and the nuclear localization signal (NLS) region in this order. Experiments with extensive deletion mutants indicated that just one deletion out of the four regions irs not enough to delete the TLS binding, although combinatorial deletion of zinc finger with other three regions almost wiped off the lncRNA3 binding. Remarkably, each of four regions alone has no binding to TLS, either. Collectively, RGG2, zinc finger, RGG3, and NLS all are essential for binding to lncRNA3, but are required to work synergistically for full binding. These data indicate that dynamic assembly of RNA-binding domain works for action of lncRNAs and possibly has allosteric effect on intrinsically disordered region (IDR) of N-terminus of TLS, implying relation of RNA-binding with phase separation and the resultant precipitation.
| Published in | Biomedical Sciences (Volume 10, Issue 2) |
| DOI | 10.11648/j.bs.20241002.13 |
| Page(s) | 30-43 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2024. Published by Science Publishing Group |
TLS/FUS, pncRNA-D, GST-TLS, Long Noncoding RNA, Phase Separation, Intrinsically Disordered Region, IDP
2.1. Antibodies and Reagents
2.2. Affinity Purification of Bacterially-Expressed GST-TLS
2.3. RNA Binding Assay
2.4. Phase Separation and Precipitation Assay of GST-TLS
2.5. Protein Analysis
2.6. RNA Sequences Used in the Experiments
3.1. LncRNAs with Preferential Interaction with TLS and Other Proteins
3.2. Extensive Mapping of RNA-Binding Ability on the Fragment 4 of TLS
3.3. Dissection of lncRNA 3 into Four Fragments and Identification of TLS Binding Site
3.4. Poly (G) RNA Promotes Precipitation of TLS
| [1] | Wang, X., et al., Induced ncRNAs allosterically modify RNA-binding proteins in cis to inhibit transcription. Nature, 2008. 454(7200): p. 126-30 |
| [2] | Vance, C., et al., Mutations in FUS, an RNA processing protein, cause familial amyotrophic lateral sclerosis type 6. Science, 2009. 323(5918): p. 1208-11 |
| [3] | Kwiatkowski, T. J., Jr., et al., Mutations in the FUS/TLS gene on chromosome 16 cause familial amyotrophic lateral sclerosis. Science, 2009. 323(5918): p. 1205-8 |
| [4] |
Lagier-Tourenne, C. and D. W. Cleveland, Rethinking ALS: the FUS about TDP-43. Cell, 2009. 136(6): p. 1001-4
https://doi.org/S0092-8674(09)00263-3[pii]10.1016/j.cell.2009.03.006 |
| [5] | Da Cruz, S. and D. W. Cleveland, Understanding the role of TDP-43 and FUS/TLS in ALS and beyond. Current Opinion in Neurobiology, 2011. 21(6): p. 904-919 |
| [6] | Taylor, J. P., R. H. Brown Jr, and D. W. Cleveland, Decoding ALS: from genes to mechanism. Nature, 2016. 539(7628): p. 197-206 |
| [7] | Van Langenhove, T., et al., Genetic contribution of FUS to frontotemporal lobar degeneration. Neurology, 2010. 74(5): p. 366-71 |
| [8] | Broustal, O., et al., FUS mutations in frontotemporal lobar degeneration with amyotrophic lateral sclerosis. Journal of Alzheimer's disease: JAD, 2010. 22(3): p. 765-9. |
| [9] | Martinelli, I., et al., G507D mutation in FUS gene causes familial amyotrophic lateral sclerosis with a specific genotype-phenotype correlation. Neurobiology of aging, 2022. 118: p. 124-128 |
| [10] | Zou, H., et al., The function of FUS in neurodevelopment revealed by the brain and spinal cord organoids. Molecular and Cellular Neuroscience, 2022. 123: p. 103771 |
| [11] | Korobeynikov, V. A., et al., Antisense oligonucleotide silencing of FUS expression as a therapeutic approach in amyotrophic lateral sclerosis. Nature Medicine, 2022. 28(1): p. 104-116 |
| [12] | Kurokawa, R., ed. Long Noncoding RNAs. 2015, Springer. 257. |
| [13] | Lipovich, L., et al., Developmental changes in the transcriptome of human cerebral cortex tissue: long noncoding RNA transcripts. Cerebral cortex, 2014. 24(6): p. 1451-9 |
| [14] | Derrien, T., et al., The GENCODE v7 catalog of human long noncoding RNAs: analysis of their gene structure, evolution, and expression. Genome research, 2012. 22(9): p. 1775-89 |
| [15] | Carninci, P., et al., The transcriptional landscape of the mammalian genome. Science, 2005. 309(5740): p. 1559-63 |
| [16] | Khalil, A. M., et al., Many human large intergenic noncoding RNAs associate with chromatin-modifying complexes and affect gene expression. Proc Natl Acad Sci U S A, 2009. 106(28): p. 11667-72 |
| [17] | Necsulea, A., et al., The evolution of lncRNA repertoires and expression patterns in tetrapods. Nature, 2014. 505(7485): p. 635-40 |
| [18] | Chi, K. R., Finding function in mystery transcripts. Nature, 2016. 529(7586): p. 423-425 |
| [19] | Kurokawa, R., Generation of Functional Long Noncoding RNA Through Transcription and Natural Selection, in Regulatory RNAs. 2012, Springer. p. 151-174. |
| [20] | Djebali, S., et al., Landscape of transcription in human cells. Nature, 2012. 489(7414): p. 101-8 |
| [21] | Hon, C.-C., et al., An atlas of human long non-coding RNAs with accurate 5′ ends. Nature, 2017. 543(7644): p. 199-204 |
| [22] | Kurokawa, R., Long noncoding RNA as a regulator for transcription. Prog Mol Subcell Biol, 2011. 51: p. 29-41 |
| [23] | Kurokawa, R., Promoter-associated long noncoding RNAs repress transcription through a RNA binding protein TLS. Advances in experimental medicine and biology, 2011. 722: p. 196-208 |
| [24] | Kurokawa, R., Initiation of Transcription Generates Divergence of Long Noncoding RNAs, in Long Noncoding RNAs. 2015, Springer. p. 69-91. |
| [25] | Aguilar, R., et al., Targeting Xist with compounds that disrupt RNA structure and X inactivation. Nature, 2022. 604(7904): p. 160-166 |
| [26] | Yang, F., et al., Shape of promoter antisense RNAs regulates ligand-induced transcription activation. Nature, 2021. 595(7867): p. 444-449 |
| [27] | Allou, L., et al., Non-coding deletions identify Maenli lncRNA as a limb-specific En1 regulator. Nature, 2021. 592(7852): p. 93-98 |
| [28] | Kurokawa, R., M. G. Rosenfeld, and C. K. Glass, Transcriptional regulation through noncoding RNAs and epigenetic modifications. RNA Biol, 2009. 6(3): p. 233-6 |
| [29] | Cech, T. R. and J. A. Steitz, The noncoding RNA revolution-trashing old rules to forge new ones. Cell, 2014. 157(1): p. 77-94 |
| [30] | Liu, S. J., et al., CRISPRi-based genome-scale identification of functional long noncoding RNA loci in human cells. Science, 2017. 355(6320): p. eaah7111 |
| [31] | Cai, Z., et al., RIC-seq for global in situ profiling of RNA–RNA spatial interactions. Nature, 2020. 582(7812): p. 432-437 |
| [32] | Elguindy, M. M. and J. T. Mendell, NORAD-induced Pumilio phase separation is required for genome stability. Nature, 2021. 595(7866): p. 303-308 |
| [33] | Quinodoz, S. A., et al., RNA promotes the formation of spatial compartments in the nucleus. Cell, 2021. 184(23): p. 5775-5790. e30 |
| [34] | Chong, P. A., R. M. Vernon, and J. D. Forman-Kay, RGG/RG Motif Regions in RNA Binding and Phase Separation. Journal of molecular biology, 2018. 430(23): p. 4650-4665 |
| [35] | Corley, M., M. C. Burns, and G. W. Yeo, How RNA-Binding Proteins Interact with RNA: Molecules and Mechanisms. Molecular Cell, 2020. 78(1): p. 9-29 |
| [36] | Lunde, B. M., C. Moore, and G. Varani, RNA-binding proteins: modular design for efficient function. Nature reviews. Molecular cell biology, 2007. 8(6): p. 479-90 |
| [37] | Clery, A., M. Blatter, and F. H. Allain, RNA recognition motifs: boring? Not quite. Current opinion in structural biology, 2008. 18(3): p. 290-8 |
| [38] |
Linder, P. and E. Jankowsky, From unwinding to clamping - the DEAD box RNA helicase family. Nature reviews. Molecular cell biology, 2011. 12(8): p. 505-16 https://doi.org/10.1038/nrm3154 |
| [39] | Ramakrishnan, V., The ribosome emerges from a black box. Cell, 2014. 159(5): p. 979-984 |
| [40] | Steitz, T. A., A structural understanding of the dynamic ribosome machine. Nature reviews. Molecular cell biology, 2008. 9(3): p. 242-53 |
| [41] | Behrmann, E., et al., Structural snapshots of actively translating human ribosomes. Cell, 2015. 161(4): p. 845-57 |
| [42] | Papasaikas, P. and J. Valcarcel, The Spliceosome: The Ultimate RNA Chaperone and Sculptor. Trends in biochemical sciences, 2016. 41(1): p. 33-45 |
| [43] | Plaschka, C., P. C. Lin, and K. Nagai, Structure of a pre-catalytic spliceosome. Nature, 2017. 546(7660): p. 617-621 |
| [44] | Beckmann, B. M., et al., The RNA-binding proteomes from yeast to man harbour conserved enigmRBPs. Nature Communications, 2015. 6(1): p. 10127 |
| [45] | Xu, Y., et al., ERα is an RNA-binding protein sustaining tumor cell survival and drug resistance. Cell, 2021. 184(20): p. 5215-5229. e17 |
| [46] | Wright, P. E. and H. J. Dyson, Intrinsically unstructured proteins: re-assessing the protein structure-function paradigm. Journal of molecular biology, 1999. 293(2): p. 321-31 |
| [47] | Han, T. W., et al., Cell-free formation of RNA granules: bound RNAs identify features and components of cellular assemblies. Cell, 2012. 149(4): p. 768-79 |
| [48] | Ozdilek, B. A., et al., Intrinsically disordered RGG/RG domains mediate degenerate specificity in RNA binding. Nucleic acids research, 2017. 45(13): p. 7984-7996 |
| [49] | Lin, Y., S. L. Currie, and M. K. Rosen, Intrinsically disordered sequences enable modulation of protein phase separation through distributed tyrosine motifs. Journal of Biological Chemistry, 2017. 292(46): p. 19110-19120 |
| [50] | Balcerak, A., et al., RNA-protein interactions: disorder, moonlighting and junk contribute to eukaryotic complexity. Open biology, 2019. 9(6): p. 190096 |
| [51] | Zeke, A., et al., Deep structural insights into RNA-binding disordered protein regions. WIREs RNA, 2022. 13(5): p. e1714 |
| [52] | Wright, P. E. and H. J. Dyson, Intrinsically disordered proteins in cellular signalling and regulation. Nat Rev Mol Cell Biol, 2015. 16(1): p. 18-29 |
| [53] | Castello, A., et al., Comprehensive Identification of RNA-Binding Domains in Human Cells. Molecular Cell, 2016. 63(4): p. 696-710 |
| [54] | Kwon, S. C., et al., The RNA-binding protein repertoire of embryonic stem cells. Nature structural & molecular biology, 2013. 20(9): p. 1122-30 |
| [55] | Liepelt, A., et al., Identification of RNA-binding Proteins in Macrophages by Interactome Capture. Molecular & cellular proteomics: MCP, 2016. 15(8): p. 2699-714 |
| [56] | Loughlin, F. E., et al., The Solution Structure of FUS Bound to RNA Reveals a Bipartite Mode of RNA Recognition with Both Sequence and Shape Specificity. Molecular cell, 2019. 73(3): p. 490-504 e6 |
| [57] | Ueda, N., R. Yoneda, and R. Kurokawa, Identification of Long Noncoding RNA Recognized by RNA-Binding Protein TLS/FUS: Purification of RNAs by Affinity Chromatography of GST-TLS. Biomedical Sciences, 2022. 8(4): p. 144-156 |
| [58] |
Du, K., et al., TLS and PRMT1 synergistically coactivate transcription at the survivin promoter through TLS arginine methylation. Biochem Biophys Res Commun, 2011. 404(4): p. 991-6
https://doi.org/S0006-291X(10)02335-1[pii]10.1016/j.bbrc.2010.12.097 |
| [59] | Cui, W., et al., Arginine methylation of translocated in liposarcoma (TLS) inhibits its binding to long noncoding RNA, abrogating TLS-mediated repression of CBP/p300 activity. The Journal of Biological Chemistry, 2018. 293(28): p. 10937-10948 |
| [60] | Ueda, N. and R. Kurokawa, Affinity Profiles Categorize RNA-Binding Proteins into Distinctive Groups. Biomedical Sciences, 2018. 4(3): p. 24-31 |
| [61] | Yoneda, R., et al., The binding specificity of Translocated in LipoSarcoma/FUsed in Sarcoma with lncRNA transcribed from the promoter region of cyclin D1. Cell & bioscience, 2016. 6: p. 4 |
| [62] | Yoneda, R., et al., Long noncoding RNA pncRNA-D reduces cyclin D1 gene expression and arrests cell cycle through RNA m(6)A modification. The Journal of biological chemistry, 2020. 295(17): p. 5626-5639 |
| [63] | Yoneda, R., N. Ueda, and R. Kurokawa, m6A Modified Short RNA Fragments Inhibit Cytoplasmic TLS/FUS Aggregation Induced by Hyperosmotic Stress. International Journal of Molecular Sciences, 2021. 22(20): p. 11014 |
| [64] | Ishiguro, A., et al., ALS-linked FUS mutations dysregulate G-quadruplex-dependent liquid-liquid phase separation and liquid-to-solid transition. J Biol Chem, 2021. 297(5): p. 101284 |
| [65] | Gallego-Iradi, M. C., et al., Subcellular Localization of Matrin 3 Containing Mutations Associated with ALS and Distal Myopathy. PLoS One, 2015. 10(11): p. e0142144 |
| [66] | Boehringer, A., et al., ALS Associated Mutations in Matrin 3 Alter Protein-Protein Interactions and Impede mRNA Nuclear Export. Sci Rep, 2017. 7(1): p. 14529 |
| [67] | Abdelmohsen, K. and M. Gorospe, RNA-binding protein nucleolin in disease. RNA Biol, 2012. 9(6): p. 799-808 |
| [68] | Peggion, C., et al., Nucleolin Rescues TDP-43 Toxicity in Yeast and Human Cell Models. Front Cell Neurosci, 2021. 15: p. 625665 |
| [69] | Hoell, J. I., et al., RNA targets of wild-type and mutant FET family proteins. Nature structural & molecular biology, 2011. 18(12): p. 1428-31 |
| [70] | Wang, X., J. C. Schwartz, and T. R. Cech, Nucleic acid-binding specificity of human FUS protein. Nucleic Acids Research, 2015. 43(15): p. 7535-43 |
| [71] | Tejedor, A. R., et al., Protein structural transitions critically transform the network connectivity and viscoelasticity of RNA-binding protein condensates but RNA can prevent it. Nature Communications, 2022. 13(1): p. 5717 |
| [72] | Du, S., et al., A nerve injury–specific long noncoding RNA promotes neuropathic pain by increasing Ccl2 expression. The Journal of Clinical Investigation, 2022. 132(13) |
| [73] | Luo, X.-J., et al., LncRNA TMPO-AS1 promotes esophageal squamous cell carcinoma progression by forming biomolecular condensates with FUS and p300 to regulate TMPO transcription. Experimental & Molecular Medicine, 2022. 54(6): p. 834-847 |
| [74] | Xing, C.-Y., et al., LINC00313 facilitates osteosarcoma carcinogenesis and metastasis through enhancing EZH2 mRNA stability and EZH2-mediated silence of PTEN expression. Cellular and Molecular Life Sciences, 2022. 79(7): p. 382 |
| [75] | Roeder, R. G., The complexities of eukaryotic transcription initiation: regulation of preinitiation complex assembly. Trends Biochem Sci, 1991. 16(11): p. 402-8. |
| [76] | Roeder, R. G., Lasker Basic Medical Research Award. The eukaryotic transcriptional machinery: complexities and mechanisms unforeseen. Nature medicine, 2003. 9(10): p. 1239-44 |
| [77] | Glass, C. K. and M. G. Rosenfeld, The coregulator exchange in transcriptional functions of nuclear receptors. Genes Dev, 2000. 14(2): p. 121-41. |
| [78] | Rosenfeld, M. G., V. V. Lunyak, and C. K. Glass, Sensors and signals: a coactivator/corepressor/epigenetic code for integrating signal-dependent programs of transcriptional response. Genes & development, 2006. 20(11): p. 1405-1428. |
| [79] | Luisi, B. F., et al., Crystallographic analysis of the interaction of the glucocorticoid receptor with DNA. Nature, 1991. 352(6335): p. 497-505 |
| [80] | Glass, C. K., et al., Mechanisms of transcriptional activation by retinoic acid receptors. Biochem Soc Trans, 1997. 25(2): p. 602-5. |
| [81] | Kurokawa, R., et al., Polarity-specific activities of retinoic acid receptors determined by a co-repressor. Nature, 1995. 377(6548): p. 451-4 |
| [82] | Kurokawa, R., et al., Differential orientations of the DNA-binding domain and carboxy-terminal dimerization interface regulate binding site selection by nuclear receptor heterodimers. Genes Dev, 1993. 7(7B): p. 1423-35. |
| [83] | Dreyfuss, G., V. N. Kim, and N. Kataoka, Messenger-RNA-binding proteins and the messages they carry. Nature reviews. Molecular cell biology, 2002. 3(3): p. 195-205 |
| [84] | Hudson, W. H. and E. A. Ortlund, The structure, function and evolution of proteins that bind DNA and RNA. Nature reviews. Molecular cell biology, 2014. 15(11): p. 749-60 |
| [85] | Hentze, M. W., et al., A brave new world of RNA-binding proteins. Nature reviews. Molecular cell biology, 2018. 19(5): p. 327-341 |
| [86] | Hasegawa, Y., et al., The matrix protein hnRNP U is required for chromosomal localization of Xist RNA. Developmental cell, 2010. 19(3): p. 469-76 |
| [87] | Lambert, N., et al., RNA Bind-n-Seq: quantitative assessment of the sequence and structural binding specificity of RNA binding proteins. Molecular Cell, 2014. 54(5): p. 887-900 |
| [88] | Zhou, Q., et al., Global pairwise RNA interaction landscapes reveal core features of protein recognition. Nature communications, 2018. 9(1): p. 2511 |
| [89] | Jankowsky, E. and M. E. Harris, Specificity and nonspecificity in RNA-protein interactions. Nature reviews. Molecular cell biology, 2015. 16(9): p. 533-44 |
| [90] | Lagier-Tourenne, C., et al., Divergent roles of ALS-linked proteins FUS/TLS and TDP-43 intersect in processing long pre-mRNAs. Nat Neurosci, 2012. 15(11): p. 1488-97 |
| [91] | Umar, M. I., C. Y. Chan, and C. K. Kwok, Development of RNA G-quadruplex (rG4)-targeting L-RNA aptamers by rG4-SELEX. Nat Protoc, 2022. 17(6): p. 1385-1414 |
| [92] | Yang, D., G-Quadruplex DNA and RNA. Methods Mol Biol, 2019. 2035: p. 1-24 |
| [93] | Lyu, J., et al., Genome-wide mapping of G-quadruplex structures with CUT&Tag. Nucleic Acids Res, 2022. 50(3): p. e13 |
| [94] | Wang, S. K., Y. Wu, and T. M. Ou, RNA G-Quadruplex: The New Potential Targets for Therapy. Curr Top Med Chem, 2015. 15(19): p. 1947-56 |
| [95] | Grossman, M., et al., Frontotemporal lobar degeneration. Nat Rev Dis Primers, 2023. 9(1): p. 40 |
| [96] | Shi, Y., et al., Structure-based classification of tauopathies. Nature, 2021. 598(7880): p. 359-363 |
| [97] | Arseni, D., et al., TDP-43 forms amyloid filaments with a distinct fold in type A FTLD-TDP. Nature, 2023. 620(7975): p. 898-903 |
| [98] | Neumann, M., et al., A new subtype of frontotemporal lobar degeneration with FUS pathology. Brain, 2009. 132(Pt 11): p. 2922-31 |
| [99] | Neumann, M., et al., Abundant FUS-immunoreactive pathology in neuronal intermediate filament inclusion disease. Acta Neuropathol, 2009. 118(5): p. 605-16 |
| [100] | Munoz, D. G., et al., FUS pathology in basophilic inclusion body disease. Acta Neuropathol, 2009. 118(5): p. 617-27 |
| [101] | Tetter, S., et al., TAF15 amyloid filaments in frontotemporal lobar degeneration. Nature, 2024. 625(7994): p. 345-351 |
| [102] | Patel, A., et al., A Liquid-to-Solid Phase Transition of the ALS Protein FUS Accelerated by Disease Mutation. Cell, 2015. 162(5): p. 1066-77 |
| [103] | Szewczyk, B., et al., FUS ALS neurons activate major stress pathways and reduce translation as an early protective mechanism against neurodegeneration. Cell Rep, 2023. 42(2): p. 112025 |
| [104] | Corcia, P., et al., Pre-symptomatic diagnosis in ALS. Rev Neurol (Paris), 2020. 176(3): p. 166-169 |
| [105] | Van Daele, S. H., et al., Genetic variability in sporadic amyotrophic lateral sclerosis. Brain, 2023. 146(9): p. 3760-3769 |
| [106] | Tyzack, G. E., et al., Widespread FUS mislocalization is a molecular hallmark of amyotrophic lateral sclerosis. Brain: a journal of neurology, 2019 |
| [107] | Tsai, Y. L., Y. C. Mu, and J. L. Manley, Nuclear RNA transcript levels modulate nucleocytoplasmic distribution of ALS/FTD-associated protein FUS. Sci Rep, 2022. 12(1): p. 8180 |
| [108] | Kato, M., et al., Cell-free formation of RNA granules: low complexity sequence domains form dynamic fibers within hydrogels. Cell, 2012. 149(4): p. 753-67 |
| [109] | Kurokawa, R. and T. Bando, Three-Dimensional Structure of RNA-Binding Protein TLS Co-Crystallized with Biotinylated Isoxazole. Biomedical Sciences, 2016. 2: p. 1-10 |
| [110] | Maharana, S., et al., RNA buffers the phase separation behavior of prion-like RNA binding proteins. Science, 2018. 360(6391): p. 918-921 |
| [111] | Zhang, Y., et al., G-quadruplex structures trigger RNA phase separation. Nucleic Acids Res, 2019. 47(22): p. 11746-11754 |
| [112] | Papageorgiou, A. C., et al., Recognition and coacervation of G-quadruplexes by a multifunctional disordered region in RECQ4 helicase. Nat Commun, 2023. 14(1): p. 6751 |
| [113] | Wang, W., et al., G-quadruplexes promote the motility in MAZ phase-separated condensates to activate CCND1 expression and contribute to hepatocarcinogenesis. Nat Commun, 2024. 15(1): p. 1045 |
| [114] | Subramanian, M., et al., G-quadruplex RNA structure as a signal for neurite mRNA targeting. EMBO Rep, 2011. 12(7): p. 697-704 |
| [115] | Ueda, N., et al., Biotin-Lys-His Blocks Aggregation of RNA-binding Protein TLS, a Cause of Amyotrophic Lateral Sclerosis. Biomedical Sciences, 2017. 3(4): p. 67-77 |
| [116] | Hamad, N., et al., Non-coding RNA suppresses FUS aggregation caused by mechanistic shear stress on pipetting in a sequence-dependent manner. Scientific reports, 2021. 11(1): p. 9523 |
APA Style
Ueda, N., Yoneda, R., Kurokawa, R. (2024). Identification of Essential Components of RNA Binding Domain of TLS/FUS. Biomedical Sciences, 10(2), 30-43. https://doi.org/10.11648/j.bs.20241002.13
ACS Style
Ueda, N.; Yoneda, R.; Kurokawa, R. Identification of Essential Components of RNA Binding Domain of TLS/FUS. Biomed. Sci. 2024, 10(2), 30-43. doi: 10.11648/j.bs.20241002.13
AMA Style
Ueda N, Yoneda R, Kurokawa R. Identification of Essential Components of RNA Binding Domain of TLS/FUS. Biomed Sci. 2024;10(2):30-43. doi: 10.11648/j.bs.20241002.13
@article{10.11648/j.bs.20241002.13,
author = {Naomi Ueda and Ryoma Yoneda and Riki Kurokawa},
title = {Identification of Essential Components of RNA Binding Domain of TLS/FUS
},
journal = {Biomedical Sciences},
volume = {10},
number = {2},
pages = {30-43},
doi = {10.11648/j.bs.20241002.13},
url = {https://doi.org/10.11648/j.bs.20241002.13},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.bs.20241002.13},
abstract = {TLS/FUS is RNA-binding protein having multiple functions of regulations of genes, homeostasis, and cellular growth. Recent studies show that TLS is involved in phase separation and occasionally forms precipitation related to neurodegenerative diseases like amyotrophic lateral sclerosis (ALS). RNA has been reported to suppress phase separation, droplet formation, and concomitant precipitation of TLS, suggesting that RNA is a possible candidate for ALS drug discovery. Our experiments demonstrated that a long noncoding RNA, promoter-associated noncoding RNA (pncRNA-D), specifically binds TLS and represses its phase separation and precipitation. To obtain competent drug seeds, it is essential to reveal mechanism of action of lncRNAs with specificity to TLS and inhibitory activity on phase separation and related precipitation. For this purpose, several lncRNAs (lncRNAs 1 to 6) were selected upon assays with GST-TLS binding and inhibition on the precipitation. With criteria of binding specificity for TLS, lncRNA3 has been selected for further analysis for RNA-binding ability. Initially, RNA-binding region at TLS amino acid sequence was identified from four fragments of TLS. RNA binding assay with biotinylated lncRNA3 precipitated with avidin magnetic beads indicated clearly that TLS binds the fragment 4 (373-526 aa), C-terminus end of TLS. Then, dissecting fragment 4 presents four regions, RGG2, zinc finger, RGG3, and the nuclear localization signal (NLS) region in this order. Experiments with extensive deletion mutants indicated that just one deletion out of the four regions irs not enough to delete the TLS binding, although combinatorial deletion of zinc finger with other three regions almost wiped off the lncRNA3 binding. Remarkably, each of four regions alone has no binding to TLS, either. Collectively, RGG2, zinc finger, RGG3, and NLS all are essential for binding to lncRNA3, but are required to work synergistically for full binding. These data indicate that dynamic assembly of RNA-binding domain works for action of lncRNAs and possibly has allosteric effect on intrinsically disordered region (IDR) of N-terminus of TLS, implying relation of RNA-binding with phase separation and the resultant precipitation.
},
year = {2024}
}
TY - JOUR T1 - Identification of Essential Components of RNA Binding Domain of TLS/FUS AU - Naomi Ueda AU - Ryoma Yoneda AU - Riki Kurokawa Y1 - 2024/05/10 PY - 2024 N1 - https://doi.org/10.11648/j.bs.20241002.13 DO - 10.11648/j.bs.20241002.13 T2 - Biomedical Sciences JF - Biomedical Sciences JO - Biomedical Sciences SP - 30 EP - 43 PB - Science Publishing Group SN - 2575-3932 UR - https://doi.org/10.11648/j.bs.20241002.13 AB - TLS/FUS is RNA-binding protein having multiple functions of regulations of genes, homeostasis, and cellular growth. Recent studies show that TLS is involved in phase separation and occasionally forms precipitation related to neurodegenerative diseases like amyotrophic lateral sclerosis (ALS). RNA has been reported to suppress phase separation, droplet formation, and concomitant precipitation of TLS, suggesting that RNA is a possible candidate for ALS drug discovery. Our experiments demonstrated that a long noncoding RNA, promoter-associated noncoding RNA (pncRNA-D), specifically binds TLS and represses its phase separation and precipitation. To obtain competent drug seeds, it is essential to reveal mechanism of action of lncRNAs with specificity to TLS and inhibitory activity on phase separation and related precipitation. For this purpose, several lncRNAs (lncRNAs 1 to 6) were selected upon assays with GST-TLS binding and inhibition on the precipitation. With criteria of binding specificity for TLS, lncRNA3 has been selected for further analysis for RNA-binding ability. Initially, RNA-binding region at TLS amino acid sequence was identified from four fragments of TLS. RNA binding assay with biotinylated lncRNA3 precipitated with avidin magnetic beads indicated clearly that TLS binds the fragment 4 (373-526 aa), C-terminus end of TLS. Then, dissecting fragment 4 presents four regions, RGG2, zinc finger, RGG3, and the nuclear localization signal (NLS) region in this order. Experiments with extensive deletion mutants indicated that just one deletion out of the four regions irs not enough to delete the TLS binding, although combinatorial deletion of zinc finger with other three regions almost wiped off the lncRNA3 binding. Remarkably, each of four regions alone has no binding to TLS, either. Collectively, RGG2, zinc finger, RGG3, and NLS all are essential for binding to lncRNA3, but are required to work synergistically for full binding. These data indicate that dynamic assembly of RNA-binding domain works for action of lncRNAs and possibly has allosteric effect on intrinsically disordered region (IDR) of N-terminus of TLS, implying relation of RNA-binding with phase separation and the resultant precipitation. VL - 10 IS - 2 ER -
Division of Biomedical Sciences, School of Medicine, Saitama Medical University, Saitama, Japan
Division of Biomedical Sciences, School of Medicine, Saitama Medical University, Saitama, Japan
Division of Biomedical Sciences, School of Medicine, Saitama Medical University, Saitama, Japan
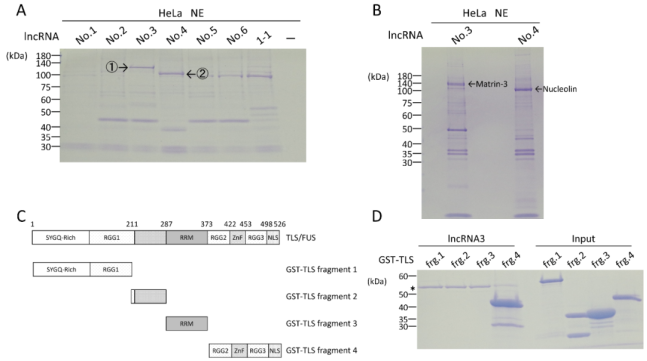
Figure 1. LncRNAs with preferential interaction with specific proteins including TLS.
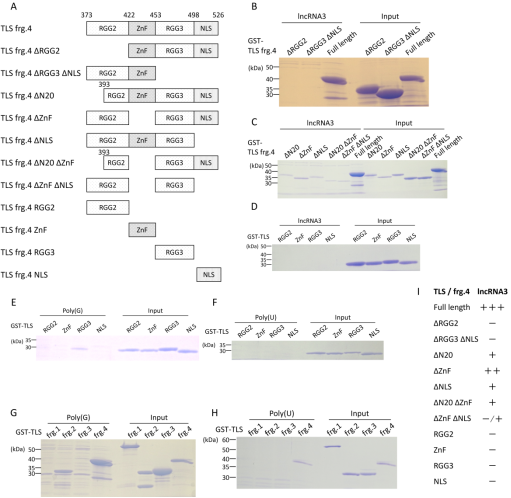
Figure 2. Extensive mapping of RNA-binding ability on the fragment 4 of TLS.
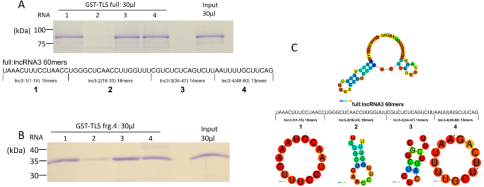
Figure 3. Dissection of lncRNA 3 into four fragments and distinctive bindings to TLS.
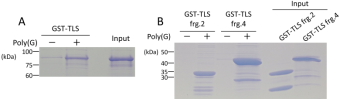
Figure 4. Poly (G) RNA promotes phase-separation induced precipitation of TLS.
Information

